Cannabidiol – CBD tăng cường độc tính tế bào của thuốc chống ung thư ở người ung thư biểu mô tế bào vảy ở đầu và cổ
- Yoon Young Go ,
- Su Ra Kim ,
- Do Yeon Kim ,
- Sung-Won Chae &
- Jae-Jun Song
Báo cáo khoa học âm lượng 10 , Bài báo số: 20622 ( 2020 ) Trích dẫn bài viết này
- 144 lượt truy cập
- 4 Altmetric
- Số liệuchi tiết
trừu tượng
Cannabidiol (CBD) có hoạt tính chống khối u. Tuy nhiên, tác dụng chống ung thư của CBD đối với ung thư biểu mô tế bào vảy ở đầu và cổ (HNSCC) vẫn chưa rõ ràng. Độc tính tế bào của CBD trên HNSCC được phân tích bằng cách sử dụng các xét nghiệm về sự sống sót của tế bào và hình thành khuẩn lạc trong ống nghiệm. RNA-seq được sử dụng để xác định cơ chế chết tế bào do CBD gây ra. Các mô hình chuột Xenograft đã được sử dụng để xác định ảnh hưởng của CBD trong cơ thể sống. Điều trị CBD làm giảm đáng kể sự di cư / xâm lấn và khả năng tồn tại của các tế bào HNSCC theo cách phụ thuộc vào liều lượng và thời gian. Mô hình xenograft chuột HNSCC cho thấy tác dụng chống khối u của CBD. Hơn nữa, điều trị kết hợp với CBD nâng cao hiệu quả của thuốc hóa trị. Quá trình apoptosis và autophagy có liên quan đến độc tính tế bào do CBD gây ra của HNSCCs. RNA-seq xác định sự giảm biểu hiện của các gen liên quan đến việc sửa chữa DNA, phân chia tế bào và tăng sinh tế bào, có liên quan đến độc tính tế bào qua trung gian CBD đối với HNSCC. Chúng tôi đã xác định CBD là một hợp chất chống ung thư tiềm năng mới cho liệu pháp đơn lẻ hoặc kết hợp HNSCC.
Giới thiệu
Ung thư biểu mô tế bào vảy ở đầu và cổ (HNSCC) là một khối u ác tính xảy ra ở các vùng lan rộng của vùng đầu và cổ, bao gồm khoang miệng và mũi, tuyến nước bọt, hầu họng và thanh quản, với một hỗn hợp không đồng nhất của các khối u ác tính đường tiêu hóa trên 1 . HNSCC được coi là loại ung thư phổ biến thứ sáu, chiếm khoảng 4% các loại ung thư trên toàn thế giới, và tỷ lệ sống sót sau 5 năm ở bệnh nhân HNSCC giai đoạn muộn là dưới 50% 2 . Hiện nay, hóa trị với cisplatin, methotrexate, và 5-fluorouracil được áp dụng cho bệnh nhân HNSCC dưới dạng đơn chất hoặc kết hợp với xạ trị để giảm diện tích khối u ban đầu trước khi phẫu thuật vật lý 3.. Các tác nhân hóa trị liệu, đặc biệt là các hợp chất bạch kim (Cisplatin), các đơn vị phân loại (Paclitaxel) và 5-Fluorouracil là những loại thuốc hiệu quả nổi tiếng trong điều trị ung thư. Tuy nhiên, việc sử dụng các đại lý hóa học trị liệu gây ra tác dụng phụ không mong muốn như mất dữ dội tóc, buồn nôn, nôn mửa, vấn đề về thận, và giảm khả năng miễn dịch với nhiễm trùng cũng như kháng thuốc 4 , 5 . Vì vậy, liệu pháp mono nhắm mục tiêu dựa trên nghiên cứu phân tử để HNSCC điều trị đã được phát triển và thử nghiệm trong nghiên cứu lâm sàng, FDA đã phê duyệt một số đại lý duy nhất cho HNSCC bao gồm Cetuximab và pembrolizumab 6 , 7. Mặc dù cetuximab được phát triển đặc biệt để nhắm mục tiêu vào thụ thể yếu tố tăng trưởng biểu bì (EGFR), được biểu hiện quá mức trong HNSCC, đáp ứng điều trị thấp và các tác dụng phụ như kháng thuốc thường hạn chế ứng dụng của nó trong hóa trị cho tất cả bệnh nhân HNSCC 6 . Do đó, các tác nhân hiệu quả hơn là cần thiết cho các chiến lược hóa trị ở bệnh nhân HNSCC.
Cannabidiol (CBD) là một trong những thành phần trong họ thực vật Cannabis sativa L. (cần sa), và nghiên cứu cơ bản đã làm sáng tỏ cấu trúc hóa học của các hợp chất CBD từ những năm 1960 8 . Cây cần sa bao gồm hai thành phần chính là CBD và tetrahydrocannabinol (THC), có hoạt tính sinh học và tác dụng chữa bệnh khác nhau đối với cơ thể con người. Ví dụ, THC đã được biết đến với đặc tính kích thích thần kinh và gây nghiện, trong khi CBD làm giảm hoạt động tâm thần của THC trong hệ thần kinh 8 . Có những lo ngại về ảnh hưởng của CBD đối với sức khỏe thể chất và tinh thần; Tuy nhiên, các nghiên cứu tiền lâm sàng cho thấy có những lợi ích y tế của CBD trong việc điều trị chứng nghiện ma túy và thuốc lá ở người 9. Do đó, CBD được coi là một thành phần dược lý an toàn của họ cannabinoid, được biết đến là có nhiều tác dụng y học, bao gồm tác dụng chống viêm và chống thiếu máu cục bộ, cộng với khả năng điều trị đau thần kinh và rối loạn co giật ở trẻ em 10 , 11 , 12 . Hiện nay, CBD được tiêu thụ rộng rãi dưới dạng chiết xuất hoặc dầu từ cây cần sa ở các nước Châu Âu và Châu Mỹ 12 .
CBD có những lợi ích tiềm năng để điều trị ung thư, bao gồm ức chế sự phát triển của khối u, hình thành mạch và di căn trong các mô hình ung thư khác nhau 13 , 14 . Cơ chế phân tử cơ bản của tác dụng chống ung thư của CBD vẫn chưa được hiểu đầy đủ, nhưng phần lớn các nghiên cứu đã chỉ ra rằng nó ức chế sự tăng sinh của tế bào ung thư thông qua tín hiệu apoptosis 15 . Ngoài ra, một trong những ưu điểm nổi tiếng nhất của CBD về tác dụng chữa bệnh của nó là giảm đau do ung thư. So với các tác dụng phụ của thuốc chống ung thư hóa trị, CBD có thể làm giảm hiệu quả buồn nôn và nôn do hóa trị liệu trong quá trình điều trị ung thư 16. Do đó, CBD có thể có một lợi thế kép cho liệu pháp điều trị ung thư. Gần đây, người ta đã báo cáo rằng cannabinoids có thể gây ra sự sinh ung thư của vi rút u nhú ở người (HPV) dương tính với HNSCC 17 . Tuy nhiên, vẫn chưa rõ tác động của CBD đối với HNSCC âm tính với HPV. Trong nghiên cứu này, chúng tôi đã tìm hiểu các cơ chế cơ bản gây chết tế bào do CBD trong các dòng tế bào HNSCC âm tính với HPV, và xác định hiệu quả hiệp đồng của việc kết hợp các tác nhân hóa trị với CBD. Ngoài ra, chúng tôi cũng cung cấp bằng chứng in vivo rằng CBD làm giảm sự phát triển của khối u ở đầu và cổ như một tác nhân gây độc tế bào hiệu quả chống lại HNSCC.
Các kết quả
Tác dụng gây độc tế bào của CBD trên tế bào HNSCC
Để điều tra tác dụng gây độc tế bào của CBD đối với tế bào HNSCC của con người, trước tiên chúng tôi đo khả năng sống của tế bào của bốn dòng tế bào HNSCC bằng xét nghiệm CCK8 sau khi xử lý với các nồng độ khác nhau của CBD (0–15 µM) trong 24 giờ. Sự giảm khả năng sống sót lớn hơn được phát hiện trong các tế bào HNSCC được nuôi cấy bằng CBD (<6 µM) so với các tế bào sừng ở miệng bình thường (HOK) của người (Hình 1 A). Ngoài ra, khả năng tồn tại của HNSCC bị CBD ức chế đáng kể theo cách phụ thuộc vào thời gian khi điều trị trong 48–72 giờ, cho thấy độc tính tế bào đáng kể của CBD như một loại thuốc chống ung thư (Hình 1B). Tương tự như kết quả xét nghiệm CCK8, kết quả xét nghiệm loại trừ màu xanh trypan cũng cho thấy cảm ứng gây chết tế bào HNSCC ở ngày thứ 2 được điều trị với các nồng độ CBD khác nhau, cho thấy CBD có tiềm năng chống ung thư trong tế bào HNSCC (Hình 1 C). Sau đó, chúng tôi điều tra xem liệu CBD có tác dụng chống di cư và chống xâm nhập lên tế bào HNSCC của người (tế bào SCC15) hay không. Đối với các xét nghiệm di cư và xâm nhập, các tế bào SCC15 được xử lý CBD và không được xử lý được cho phép chuyển gen trong các hệ thống chèn transwell và sau đó số lượng tế bào đã di cư / xâm nhập được xác định. Số lượng tế bào SCC15 được chuyển nạp giảm đáng kể khi điều trị CBD phụ thuộc vào liều lượng, so với số lượng tế bào SCC15 đối chứng trong cả hệ thống xét nghiệm di trú và xâm nhập (Hình 1D, E). Những kết quả này chứng minh rằng CBD ức chế sự di cư và xâm nhập của các tế bào HNSCC ở người và cũng có tác dụng gây độc tế bào.

Cannabidiol (CBD) gây độc tế bào ở các tế bào ung thư biểu mô tế bào vảy ở đầu và cổ (HNSCC). ( A ) Một dòng tế bào bình thường (HOK) và tế bào HNSC (FaDu, SNU899, SCC15, Hep2) được xử lý với các nồng độ khác nhau (0, 0,1, 0,5, 1, 5, 10, 15 µM) của CBD trong 24 giờ và sau đó khả năng sống của tế bào được xác định bằng xét nghiệm CCK8. EC 50 cho biết nồng độ hiệu quả đối với 50% tế bào chết. ( B ) Tế bào FaDu, SCC15 và Hep2 được xử lý với 0, 0,1, 0,5, 1, 3, 5 và 10 µM CBD. Khả năng sống sót của tế bào được đo bằng xét nghiệm CCK8 ở 24, 48 và 72 giờ. ( C) Tế bào FaDu được gieo hạt trong đĩa 24 giếng và sau đó được tiếp xúc với CBD trong mỗi môi trường tăng trưởng trong 48 giờ. Các tế bào nhuộm bằng dung dịch xanh trypan 0,4% được quan sát bằng kính hiển vi ánh sáng. Thanh chia độ đại diện cho 200 μm. ( D , E ) Các tế bào SCC15 được tiếp xúc với 0 hoặc 6 µM CBD trong 24 giờ và sau đó thêm các tế bào vào các miếng đệm xuyên thấu phía trên có phủ Matrigel hoặc không tráng phủ trong môi trường không có huyết thanh. Sau khi ủ trong 24 giờ, các tế bào đã biến nạp được nhuộm bằng dung dịch Diff Quick và sau đó được quan sát bằng kính hiển vi ánh sáng. Các ô di chuyển và xâm nhập được đếm cho mỗi nhóm. Thang chia vạch, 200 μm. Dữ liệu được biểu diễn dưới dạng giá trị trung bình và độ lệch chuẩn (SD) (n = 3); * p <0,05; ** p <0,01; và *** p <0,001 so với đối chứng tương ứng.
Tác dụng chống khối u của CBD in vivo
Tiếp theo, chúng tôi xác nhận thêm khả năng chống khối u của CBD bằng cách sử dụng mô hình chuột xenograft. Hai mô hình chuột xenograft tế bào FaDu khác nhau được tạo ra bằng cách tiêm 2 × 10 6 tế bào vào dưới da hoặc vào lưỡi của chuột khỏa thân BALB / c. Như được thể hiện trong Hình 2 A, kích thước khối u của khối u xenograft lưỡi đã giảm đáng kể ở những con chuột được điều trị bằng CBD so với những con ở những con chuột đối chứng không được điều trị. Kết quả này chỉ ra rằng CBD ức chế sự phát triển của tế bào HNSCC in vivo. Hơn nữa, sự ức chế đáng kể sự phát triển của khối u và trọng lượng đã được quan sát thấy ở những con chuột được điều trị kết hợp với cả CBD và Cisplatin (Cis) (Hình 2 B – D), có nghĩa là CBD có thể được sử dụng kết hợp với các loại thuốc chống ung thư nói chung như với tư cách là Cis.

Tác dụng chống khối u của cannabidiol (CBD) trên mô hình động vật xenograft ung thư biểu mô tế bào vảy ở đầu và cổ (HNSCC). ( A ) Tế bào FaDu (2 × 10 6 ) được tiêm vào lưỡi của chuột khỏa thân. Sau 2-3 ngày, chuột được điều trị bằng EtOH hoặc CBD (5 mg / kg) hoặc trong phúc mạc (ip). Nhuộm H&E của chuột xenograft tế bào FaDu (lưỡi) được điều trị bằng CBD hoặc EtOH sau 3 tuần được quan sát bằng kính hiển vi ánh sáng và kích thước khối u được đo. Phần trăm diện tích khối u được xác định bằng phần mềm Image J. Các thanh tỷ lệ thể hiện 200 μm. ( B ) Ô FaDu (2 × 10 6) được tiêm dưới da cho con vật. Chuột xenograft tế bào FaDu (dưới da) được điều trị bằng EtOH, CBD (5 mg / kg), Cisplatin (2,5 mg / kg) hoặc CBD (5 mg / kg) / Cisplatin (2,5 mg / kg) (CBD + Cis) sau 4 tuần đã được hy sinh và các bức ảnh của chuột và khối u được chụp bằng máy ảnh kỹ thuật số. EtOH hoặc CBD được điều trị bằng đường uống (oa) và Cisplatin được tiêm trong phúc mạc (ip). Thể tích khối u ( C ) và trọng lượng ( D ) được xác định một cách thống kê. (n = 4 chuột / nhóm và 3 hình ảnh / chuột) Dữ liệu được biểu diễn dưới dạng trung bình và SD; * p <0,05 và ** p <0,01 so với đối chứng tương ứng.
CBD tăng cường tác dụng của thuốc chống ung thư trong HNSCC
Các loại thuốc hóa trị liệu chống ung thư nói chung bao gồm Cis, 5-Fluorouracil (5-FU) và Paclitaxel (Taxol) làm giảm tỷ lệ sống sót của tế bào FaDu theo cách phụ thuộc vào liều lượng (Hình bổ sung S1 ); cụ thể, 10 µM Cis, 18 µM 5-FU, hoặc 15,2 µM Taxol trong các nghiệm thức đơn lẻ được xác định là nồng độ hiệu quả đối với tử vong 50% trong các dòng tế bào HNSCC (Hình 3 B bảng dưới). Để đánh giá xem liệu CBD có tăng cường hiệu quả của các loại thuốc chống ung thư hay không, chúng tôi đã so sánh tỷ lệ sống sót của tế bào khi điều trị đơn lẻ với một loại thuốc hóa học chống ung thư nói chung với tỷ lệ sống sót khi điều trị kết hợp với CBD (Hình 3 A). Các tấm dưới của Hình 3B cho thấy sự hiệp đồng mạnh mẽ trong các tế bào FaDu khi kết hợp thuốc có tỷ lệ CBD: Cisplatin hoặc CBD: 5-Fu và / hoặc Paclitaxel dưới 6: 5 hoặc 6:10, tương ứng. Kết hợp với 3 µM hoặc 6 µM CBD ức chế rõ rệt khả năng sống của tế bào FaDu so với liệu pháp đơn lẻ thuốc chống ung thư, cho thấy rằng một lượng Cis, 5-FU và Taxol thấp hơn đáng kể là đủ để tiêu diệt 50% tế bào HNSCC. (Hình 3 C). Để phân tích hiệu quả lâu dài của việc kết hợp CBD với thuốc chống ung thư đối với sự sống còn của tế bào FaDu được tạo dòng vô tính, các thử nghiệm hình thành khuẩn lạc đã được thực hiện. So với điều trị đơn lẻ với 5-FU, Cis hoặc Taxol, điều trị đồng thời với CBD có hiệu quả hơn đối với liệu pháp chống ung thư (Hình 3 D và Hình bổ sung S2). Tổng hợp lại, những kết quả này chỉ ra rằng sự kết hợp thuốc độc tế bào với CBD có hiệu quả hơn trong điều trị ung thư.

Cannabidiol (CBD) thể hiện hiệu quả hiệp đồng với các loại thuốc chống ung thư. ( A ) Tế bào FaDu được xử lý với các nồng độ khác nhau của Cisplatin, 5-Fluorouracil và Paclitaxel trong 24 giờ và sau đó khả năng sống của tế bào được xác định bằng xét nghiệm CCK8 để xác định giá trị EC 50 của mỗi loại thuốc. Sau bốn giờ tiền xử lý với Cisplatin, 5-Fluorouracil hoặc Paclitaxel với các tế bào FaDu được tiếp xúc với 3 hoặc 6 µM CBD trong mỗi nhóm thuốc. Tỷ lệ sống sót của tế bào được xác định bằng cách sử dụng xét nghiệm CCK8 ở 24 giờ. ( B ) Phương pháp Isobologram được sử dụng để phân tích tính cộng hưởng và / hoặc sức mạnh tổng hợp của điều trị kết hợp thuốc in vitro trong tế bào FaDu. Đường chéo mô tả sự ức chế sinh trưởng 95%. Giá trị Chỉ số Kết hợp (CI) được xác định bằng cách sử dụng 50% nồng độ ức chế (IC 50) các giá trị từ mỗi sự kết hợp thuốc, được chỉ bằng các hình tròn hoặc hình tam giác màu. Tính đối kháng (CI> 1) và sức mạnh tổng hợp (CI <1) lần lượt được chỉ ra ở trên và dưới dòng. ( C ) Điều trị đơn lẻ Cisplatin, 5-Fluorouracil hoặc Paclitaxel được so sánh với điều trị kết hợp với 3 hoặc 6 µM CBD và biểu thị giá trị EC 50 dưới dạng biểu đồ và bảng. ( D ) Tế bào FaDu được tiếp xúc với giá trị EC 50 của Cisplatin (10 µM), 5-Fluorouracil (18 µM), và Paclitaxel (15 µM) đơn lẻ hoặc kết hợp với 6 µM CBD và sau đó được nuôi cấy trong 2 tuần. Các tế bào được nhuộm bằng màu tím pha lê và được quan sát bằng kính hiển vi ánh sáng. Thang chia vạch, 200 μm. Các thanh lỗi chỉ ra giá trị trung bình và SD; * p <0,05; ** p <0,01; và *** p <0,001 so với đối chứng.
CBD gây ra apoptosis và autophagy trong các tế bào HNSCC
Chúng tôi thấy rằng CBD ức chế đáng kể sự tăng sinh của tế bào HNSCC trong nghiên cứu này. Tuy nhiên, các phản ứng tế bào do CBD gây ra trong các tế bào HNSCC, đặc biệt là tín hiệu chết của tế bào, vẫn chưa được biết. Để kiểm tra xem CBD có kích hoạt tín hiệu apoptotic trong HNSCC hay không, trước tiên chúng tôi thực hiện nhuộm Annexin V / PI của ba dòng tế bào HNSCC chưa được điều trị hoặc được điều trị bằng CBD. Phụ thuộc vào liều lượng CBD làm tăng đáng kể số lượng tế bào nhuộm màu dương tính kép Annexin V / PI so với số lượng tế bào HNSCC không được điều trị, cho thấy CBD đã kích thích quá trình apoptosis trong tế bào HNSCC (Hình 4 A). Một thí nghiệm khác, được hiển thị trong Hình 4 B, cho thấy rằng việc xử lý các tế bào SCC15 với 10 µM CBD làm giảm mức mRNA của TP53 và Bcl-2, trong khi mứcBax và caspase-3 và -9 đã tăng lên đáng kể (Hình 4 B, bảng điều khiển bên trái). Trong số các dấu hiệu apoptotic này, điều trị CBD làm giảm biểu hiện p53, có nghĩa là mức độ biểu hiện p53 thấp có thể liên quan đến quá trình bắt giữ chu kỳ tế bào, nhưng không phải là quá trình apoptosis, khi các tế bào HNSCC tiếp xúc với CBD 18 . Phân tích immunoblot cũng cho thấy rằng liều lượng CBD phụ thuộc vào việc tăng biểu hiện của PARP và -caspase 7 bị phân cắt trong các tế bào SCC15 (Hình 4 B, bảng bên phải). Tiếp theo, chúng tôi điều tra xem liệu cái chết của tế bào qua trung gian truyền tín hiệu autophagy có liên quan đến việc gây độc tế bào do CBD của HNSCCs hay không. Tăng mức độ biểu hiện của các gen đánh dấu autophagy, bao gồm Beclin –và các gen mã hóa LC3II đã được quan sát thấy trong các tế bào HNSCC được xử lý CBD (Hình 4 C, bảng điều khiển bên trái). Chúng tôi cũng phát hiện rõ ràng sự gia tăng mức protein LC3II trong các tế bào FaDu và SCC15 được xử lý CBD (Hình 4 C, bảng bên phải). Tuy nhiên, tính năng autophagy do CBD gây ra có thể có chức năng bảo vệ chống lại HNSCC. Do đó, chúng tôi đã ức chế autophagy bằng cách thêm chloroquine (CQ) trước khi điều trị với CBD vào HNSCC và sau đó thực hiện xét nghiệm CCK8 để xác định khả năng sống sót của tế bào. Điều trị bằng CQ dẫn đến sự gia tăng đáng kể khả năng sống sót của các tế bào FaDu, Hep2 và SCC15 được điều trị CBD (Hình 4D), ngụ ý rằng quá trình tự chết do CBD gây ra đã tăng cường quá trình chết tế bào trong tế bào HNSC, nhưng không phải là cơ chế bảo vệ. Nhìn chung, độc tính qua trung gian CBD đối với tế bào HNSCC liên quan đến cả cơ chế apoptosis và autophagy trong tín hiệu chết của tế bào.

Cannabidiol (CBD) gây ra cái chết của tế bào ung thư biểu mô tế bào vảy ở đầu và cổ (HNSCC) ở người thông qua quá trình apoptosis và autophagy. ( A ) Tế bào SCC15, Hep2 và FaDu được xử lý với 0, 3, 6, hoặc 10 µM CBD trong 24 giờ và nhuộm bằng Annexin V và PI. Apoptosis được phát hiện bằng phương pháp đo tế bào dòng chảy. ( B ) Tế bào SCC15 được ủ với CBD với liều lượng được chỉ định và sau đó thực hiện định lượng RT-PCR thời gian thực và phân tích Western blot. Các mức mRNA của p53, Bcl-2, Bax, caspase-3, và -9 và mức protein của PARP và caspase-7 được phân tích khi tiếp xúc với CBD trong tế bào HNSCC. ( C) Mức độ biểu hiện của các gen mã hóa Beclin- và LC3II được xác định bằng RT-PCR thời gian thực định lượng sau khi xử lý với 10 µM CBD trong 24 giờ. Mức protein LC3II được xác định trong các tế bào FaDu và SCC15 được xử lý với 0 hoặc 6 µM CBD trong 24 giờ. ( D ) Tiền xử lý các tế bào FaDu, Hep2 và SCC15 với CQ được tiếp xúc với CBD 0 hoặc 6 µM trong 24 giờ. Khả năng sống của tế bào được xác định bằng xét nghiệm CCK8. Dữ liệu được biểu diễn dưới dạng giá trị trung bình và độ lệch chuẩn (SD) (n = 3); * p <0,05 so với đối chứng tương ứng. Gel và blots có độ dài đầy đủ ở ( B ) và ( C ) được đưa vào quy trình thử nghiệm bổ sung.
Những thay đổi trên toàn bộ bộ gen được kích hoạt bởi CBD trong HNSCC: tổn thương DNA và bắt giữ chu kỳ tế bào
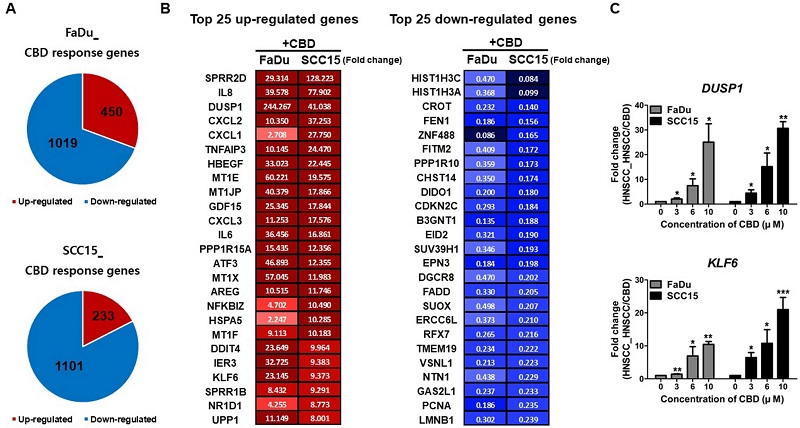
Tiếp theo, chúng tôi điều tra những thay đổi ở cấp độ toàn bộ bảng điểm được kích hoạt bởi điều trị CBD trong HNSCC. Phân tích RNA-seq đã xác định 1469 và 1334 gen trong các tế bào FaDu và SCC15, tương ứng là các gen biểu hiện khác biệt có ý nghĩa (DEGs; <hai lần) trong các tế bào HNSC được xử lý CBD. Đối với tế bào FaDu, 450 gen được điều chỉnh tăng đáng kể, trong khi 1019 gen được điều chỉnh giảm. Các tế bào SCC15 được xử lý CBD thể hiện 233 gen điều hòa đáng kể và 1101 gen điều hòa giảm (Hình 5 A). Bảng B trong Hình 5 cho thấy 25 gen quy định lên và xuống trên cùng được kết hợp trong các tế bào FaDu và SCC15. Từ các gen quy định ở cả hai dòng tế bào, chúng tôi chọn ra hai gen, DUSP1 (phosphatase đặc hiệu kép 1) và KLF6(Kruppel giống như yếu tố 6), là mục tiêu cuối cùng của tín hiệu EGFR và sau đó xác nhận sự gia tăng mức độ biểu hiện phụ thuộc vào liều lượng của cả hai gen trong HNSCC được điều trị CBD (Hình 5 C). Ngoài ra, chúng tôi đã phân tích 50 gen quy định lên và xuống hàng đầu trong các tế bào FaDu và SCC15, tương ứng (Hình bổ sung S3 ) 60 gen liên quan đến ung thư trong các tế bào FaDu và SCC15 được điều trị CBD được tìm thấy trong phân tích của chúng tôi ( SI 1 ). Đối với DEG lên và xuống được điều chỉnh bởi gấp đôi hoặc lớn hơn trong các tế bào FaDu được xử lý CBD, các thuật ngữ bản thể học gen (GO) quan trọng trong các quá trình sinh học bao gồm điều hòa tích cực của quá trình apoptosis, điều hòa tiêu cực về tăng sinh tế bào và điều chỉnh giảm quá trình sao chép DNA và sửa chữa (Hình 6A). GADD45A (bắt giữ tăng trưởng và protein alpha gây tổn thương DNA) tham gia vào quá trình sửa chữa DNA và quá trình apoptosis 19 . CDKN1A (chất ức chế kinase 1A phụ thuộc cyclin), còn được gọi là p21, có liên quan đến quá trình bắt giữ chu kỳ tế bào liên kết với tổn thương DNA 20 . Quan trọng hơn, mạnh mẽ lên điều chỉnh của GADD45A và CDKN1A biểu hiện gen đã được quan sát ở cả tế bào FaDu và SCC15 CBD xử lý (Hình. 6 B), cho thấy rằng CBD có thể gây ra apoptosis và tế bào bắt giữ chu kỳ bằng cách điều chỉnh mức GADD45A và p21 trong tế bào HNSCC. Ngoài ra, chu kỳ tế bào và các gen liên quan đến tổn thương / sửa chữa DNA như PARP1 (poly ADP-ribose polymerase 1), ATR (liên quan đến ATM và Rad3), XRCC3(Nhóm bổ sung chéo sửa chữa tia X 3), CDK5 (kinase 5 phụ thuộc cyclin) và CHEK1 (kinase điểm kiểm tra 1) cho thấy mức độ biểu hiện giảm đáng kể trong HNSCC được điều trị bằng CBD (Hình 6 B). Mức độ biểu hiện được điều chỉnh giảm của gen MCM2 (phức hợp duy trì nhiễm sắc thể nhỏ 2), PARP1 và BRCA1 (ung thư vú loại 1) trong HNSCC được điều trị bằng CBD được xác nhận bằng định lượng RT-PCR thời gian thực và sau đó so sánh với RNA- kết quả seq (Hình 6 C). Các gen biểu hiện đáng kể khác liên quan đến sửa chữa DNA, chu kỳ tế bào, tăng sinh tế bào và di chuyển tế bào trong HNSCC được điều trị CBD cũng được xác định bằng dữ liệu của
chúng tôi ( SI 2 ).

Cannabidiol (CBD) gây ra những thay đổi về bản sao lan rộng trong các tế bào ung thư biểu mô tế bào vảy ở đầu và cổ (HNSCC). ( A ) Tế bào FaDu và SCC15 được xử lý với 10 µM CBD và RNA-seq được thực hiện để điều tra các gen biểu hiện khác biệt (DEG) bằng cách xử lý CBD trên HNSCC. Số lượng gen quy định tăng và giảm phản ứng với CBD trong các tế bào FaDu và SCC15 được chỉ ra tương ứng. ( B ) 50 gen quy định lên và xuống hàng đầu ở cả FaDu và SCC15 được biểu thị bằng hình ảnh màu sắc và giá trị của sự thay đổi nếp gấp (chênh lệch gấp đôi với p <0,05). Màu sắc mô tả mức độ thay đổi nếp gấp. ( C) Các gen điều hòa đại diện mã hóa DUSP1 và KLF6 trong các tế bào FaDu và SCC15 được CBD xử lý đã được xác nhận bằng RT-PCR thời gian thực định lượng. Dữ liệu chỉ ra trung bình và SD; * p <0,05; ** p <0,01; và *** p <0,001 so với đối chứng tương ứng.

Thay đổi các quy trình sinh học bằng cách xử lý cannabidiol (CBD). ( A ) Các thuật ngữ bản thể học gen quan trọng của các quá trình sinh học liên quan từ các gen quy định lên và xuống khác biệt trong các tế bào FaDu được xử lý CBD đã được liệt kê (sự khác biệt gấp đôi với p <0,05). ( B ) Danh sách cho thấy các gen liên quan đến sự bắt giữ chu kỳ tế bào và tổn thương DNA trong dữ liệu RNA-seq. ( C ) Mức độ biểu hiện của các gen điều chỉnh giảm mã hóa MCM2, PARP1 và BRCA1 được xác nhận bằng RT-PCR thời gian thực định lượng và so sánh với kết quả của RNA-seq.
Thảo luận
HNSCC có căn nguyên do tiếp xúc với các chất gây ung thư như thuốc lá và rượu, và nhiễm vi rút gây u nhú ở người (HPV). Đối với điều trị chính, kết hợp thuốc độc tế bào và xạ trị là chiến lược điều trị duy nhất có sẵn cho bệnh nhân HNSCC 6 . Một số ít tế bào ung thư mọc lại và phát triển tại chỗ hoặc hình thành di căn xa theo ba bước sinh lý bệnh: xâm lấn, vận chuyển và tái phát 21 . Tái phát tại chỗ và di căn ung thư là những thách thức hiện nay đối với điều trị HNSCC 6. Để ức chế và tiêu diệt tế bào ung thư một cách hiệu quả, một trong những lựa chọn được đề xuất gần đây cho hóa trị liệu chống ung thư chủ yếu bao gồm cocktail thuốc chống ung thư sử dụng các hóa chất độc tế bào và kìm tế bào, cũng như liệu pháp sinh học (kháng thể, vắc xin, liệu pháp gen và liệu pháp miễn dịch, v.v.) 22. Ở đây, chúng tôi nhận thấy rằng CBD có tiềm năng chống ung thư như một loại thuốc gây độc tế bào cho HNSCC, và việc kết hợp CBD sản phẩm tự nhiên với các loại thuốc hóa trị liệu như fluoropyrimidines, các chất tương tự bạch kim hoặc các đơn vị phân loại có thể ức chế sự phát triển của tế bào ung thư nhiều hơn so với điều trị bằng thuốc đơn lẻ. Thuốc độc tế bào để hóa trị có thể có một số tác dụng phụ đối với bệnh nhân ung thư, bao gồm mệt mỏi, buồn nôn, rụng tóc và tiêu chảy vì làm tổn thương các tế bào bình thường. Tuy nhiên, CBD có thể tạo ra tác dụng giảm nhẹ và ít độc hại hơn đối với các tế bào bình thường như một hợp chất tự nhiên, và chúng tôi cũng tiết lộ rằng CBD gây ra độc tính tế bào ở HNSCC ở người, nhưng không gây ra ở các tế bào sừng thông thường ở miệng. Vì thế, điều trị kết hợp với CBD có thể làm giảm hiệu quả các tác dụng phụ khó chịu ở bệnh nhân ung thư bằng cách giảm sử dụng các loại thuốc hóa trị liệu độc hại trong quá trình điều trị ung thư. Trong bối cảnh tiềm năng điều trị ung thư của CBD để sử dụng trong lâm sàng, một nghiên cứu trong tương lai nên so sánh hiệu quả của CBD giữa HNSCC tiên phát và tiên tiến / tái phát.
Người ta đã biết rằng CBD kích hoạt quá trình chết tế bào qua trung gian apoptosis trong một số dòng tế bào ung thư như tế bào ung thư vú, đại trực tràng, bệnh bạch cầu và khối u tuyến tụy 23 , 24 , 25 , 26. Nhất quán, chúng tôi cũng quan sát thấy quá trình chết rụng phụ thuộc vào liều qua trung gian CBD trong HNSCC. Hơn nữa, chúng tôi phát hiện ra rằng điều trị với CBD trong HNSCC làm tăng mức độ PARP1 và LC3 II bị phân cắt, có nghĩa là tác dụng gây độc tế bào của CBD trong HNSCC có liên quan đến quá trình tự chết cũng như tự chết. Trong nghiên cứu của chúng tôi, việc ức chế autophagy đã ngăn chặn quá trình chết tế bào do CBD gây ra ở HNSCC, cho thấy rằng autophagy do CBD có liên quan rõ ràng đến cơ chế chết của tế bào, nhưng không có vai trò bảo vệ của autophagy đối với HNSCC. Một số nhà nghiên cứu trước đây đã chứng minh rằng tín hiệu autophagy là ngược dòng của quá trình apoptosis trong cơ chế chết tế bào gây ra bởi CBD 27 , 28. Chúng tôi không tìm hiểu cơ chế chết tế bào nào kiểm soát quá trình chết tế bào do CBD gây ra, nhưng đã chứng minh rõ ràng rằng cả quá trình apoptosis và autophagy đều có liên quan đến cơ chế chết tế bào qua trung gian CBD trong HNSCC.
p53 thường bị bất hoạt bởi các đột biến trong hầu hết các trường hợp HNSCC, và do đó không có khả năng tạo điều kiện bắt đầu quá trình apoptosis. Trong nghiên cứu của chúng tôi, gen p53 đã được điều chỉnh giảm bởi xử lý CBD được xác định bởi RNA-seq và PCR. Những kết quả này có thể thể hiện vai trò phi chức năng của p53 đối với quá trình chết rụng qua trung gian CBD trong HNSCC. Tuy nhiên, PARP1 phân cắt đã được quan sát thấy phụ thuộc vào liều lượng CBD trong HNSCC, có thể gây ra apoptosis nội bào.
Erener và cộng sự. 29 gợi ý về vai trò không phụ thuộc vào quá trình apoptosis của sự phân cắt caspase 7 và PARP1, giới thiệu cái nhìn mới về tín hiệu viêm. Sự gia tăng caspase 7 phân cắt và PARP1 được quan sát trong nghiên cứu này có thể là bằng chứng của quá trình apoptosis, nhưng nó cũng có thể tạo điều kiện cho chất nhiễm sắc mở các gen liên quan đến tín hiệu NF-kB, do đó thúc đẩy sự biểu hiện của các gen tiền viêm. Thật vậy, các gen phản ứng viêm mã hóa IL-8, IL-6R, NFKBIZ, IL-1R và PTGS2 đã được điều chỉnh đáng kể trong dữ liệu RNA-seq ( SI 3 ) của chúng tôi gợi ý rằng sự phân chia caspase 7 và PARP1 do CBD gây ra có thể kích thích viêm tế bào phản hồi trong HNSCC.
DUSP1 có thể dephosphorylate hóa cả gốc tyrosine và threonine như một kinase phosphatase đặc hiệu kép, là chất điều hòa âm tính đã biết của tín hiệu MAPK (mitogen hoạt hóa protein kinase) do EGFR khởi xướng. DUSP1 có khả năng bất hoạt các đồng dạng MAPK trong tín hiệu Ras / ERK gây ung thư thông qua hoạt động của quá trình khử phosphoryl hóa đặc hiệu kép 30 . Kích hoạt con đường MAPK qua trung gian Ras thúc đẩy chu kỳ tế bào và sự tăng sinh, dẫn đến các khối u ác tính 31 . Các nghiên cứu gần đây coi DUSP1 như một phân tử đích chống ung thư để điều chỉnh quá trình tự chết và tín hiệu autophagy của con đường MAPK được kích hoạt quá mức trong các tế bào ung thư 32 , 33 . Chúng tôi cũng nhận thấy rằng DUSP1là một trong những gen được điều chỉnh nhiều nhất bởi RNA-seq khi điều trị CBD trên HNSCC, và đã xác nhận mức độ biểu hiện gia tăng của nó trong nghiên cứu này. Cơ chế chính xác của CBD trong HNSCC vẫn chưa được biết, nhưng kết quả của chúng tôi có thể cho thấy CBD ngăn chặn hiệu quả sự tăng sinh và phát triển của tế bào HNSCC bằng cách gây ra hoạt động apoptotic và autophagy của DUSP1.
Hơn nữa, mức độ biểu hiện tăng của KLF6 cũng được khảo sát trong nghiên cứu của chúng tôi. Sự biểu hiện quá mức của KLF6 đã được biết là gây ra quá trình apoptosis và ức chế sự tăng sinh và di chuyển của tế bào trong các tế bào ung thư khác nhau; một nghiên cứu cho thấy chức năng ức chế khối u của KLF6 trong các tế bào ung thư miệng 34 . Ngoài ra, KLF6 có liên quan đến việc bất hoạt đường truyền tín hiệu qua trung gian PI3K do EGFR khởi xướng bằng cách ức chế kích hoạt AKT và mTOR. Trên thực tế, EGFR được biểu hiện quá mức trong HNSCC và được coi là một mục tiêu điều trị quan trọng, nhưng một số liệu pháp dựa trên kháng EGFR không có hiệu quả 35. Một trong những vấn đề chính của liệu pháp điều trị dựa trên EGFR trong tế bào ung thư là sự kích hoạt cơ bản của tín hiệu gây ung thư qua trung gian EGFR (MAPK, PI3K / AKT, JAK / STAT và con đường AKT / NF-kB) bằng đột biến EGFR, do đó dẫn đến sự phát triển của kháng điều trị 36 . Do đó, CBD có tiềm năng lớn như một loại thuốc chống ung thư cho HNSCC, không chỉ do tác dụng gây độc tế bào của nó bằng cách gây ra căng thẳng nhiễm độc gen, mà còn vì nó nhắm mục tiêu cụ thể đến các chất trung gian đầu cuối EGFR bằng cách điều chỉnh biểu hiện DUSP1 và KLF6. Kết quả của chúng tôi từ RNA-seq với các phát hiện in vitro và in vivo cho thấy rằng CBD có thể gây ra sự biểu hiện của DUSP1 và KLF6, sau khi ức chế khối u HNSCC thông qua sự kích hoạt phụ thuộc DUSP1 và / hoặc KLF6 của quá trình chết rụng và tín hiệu tế bào autophagy trên HNSCC.
Điều thú vị là các tế bào FaDu được xử lý bằng CBD thể hiện quá trình sao chép và sửa chữa DNA được điều chỉnh giảm đáng kể, xác nhận mức độ biểu hiện không được kiểm soát của các gen liên quan như MCM2 , PARP1 và BRCA1 . BRCA1 và PARP1 đóng một vai trò quan trọng trong việc sửa chữa các đứt gãy DNA sợi kép và sợi đơn, lần lượt là 37 . Ngoài ra, MCM2 là một vi khuẩn khởi tạo sao chép bắt đầu sao chép DNA thích hợp 38 . Nếu sự tổn thương và sao chép DNA trở nên không kiểm soát được do thiếu các yếu tố quan trọng liên quan đến quá trình sửa chữa và sao chép DNA, thì sự phá hủy DNA sẽ tăng lên, sau đó gây ra quá trình apoptosis ở các tế bào ung thư 5. Vẫn chưa rõ CBD can thiệp vào quá trình sao chép và sửa chữa DNA như thế nào, nhưng nghiên cứu của chúng tôi đã đề xuất một cơ chế hoạt động có thể có của CBD trên HNSCC: Tổn thương DNA do CBD gây ra thông qua việc điều chỉnh MCM2, PARP1 hoặc BRCA1 có thể được cho là gây ra quá trình chết rụng và CBD cũng kích hoạt KLF6 sau đó, có thể thúc đẩy biểu hiện CDKN1A (p21) trong tế bào. Sự biểu hiện protein p21 tăng lên do đó gây ra quá trình bắt giữ chu kỳ tế bào thông qua việc ức chế cyclin D và kích thích biểu hiện GADD45A / B (Hình 7 ).

Trình bày sơ đồ các cơ chế chống ung thư được đề xuất bằng cách điều trị CBD trên HMEECs.
Kết luận, nghiên cứu của chúng tôi đã xác định tiềm năng chống khối u của CBD. Ngoài ra, điều trị CBD đơn lẻ hoặc đồng điều trị với các tác nhân hóa trị liệu thúc đẩy quá trình chết tế bào HNSCC cùng với quá trình apoptosis và autophagy. Do đó, nghiên cứu của chúng tôi cho thấy CBD có thể là một chất điều trị tuyệt vời chống lại HNSCC.
Nguyên liệu và phương pháp
Dòng tế bào và quy trình nuôi cấy
Tế bào sừng ở miệng của người (HOK) được sử dụng như một dòng tế bào kiểm soát chống lại HNSCC như được mô tả 4 , thu được từ ScienCell (# 2610) và được nuôi cấy với môi trường tăng trưởng tế bào sừng ở miệng có bổ sung penicilin / streptomycin (1000 U / mL, Gibco, New York Thành phố, NY, Hoa Kỳ). Các tế bào HNSCC bao gồm FaDu (KCLB-30043), Hep2 (KCLB-10023) và SNU-899 (KCLB-00899) đã được mua từ Ngân hàng Di động Hàn Quốc (KCLB, www.cellbank.snu.ac.kr) và SCC15 được lấy từ Bộ sưu tập Văn hóa Kiểu Mỹ (ATCC, Manassas, VA, USA; CRL-1623). Tế bào FaDu, Hep2 và SCC15 được nuôi cấy trong DMEM có hàm lượng glucose thấp (Lonza, Walkersville, MD, USA) với 10% huyết thanh bò thai (FBS) (Gibco), và 1000 U / mL penicilin / streptomycin (Gibco). Dòng tế bào SNU-899 được duy trì bằng môi trường RPMI-1640 (Lonza) được bổ sung 10% FBS và penicillin / streptomycin (1000 U / mL, Gibco). Tất cả các tế bào được nuôi cấy phụ ở khoảng 90% hợp lưu và duy trì ở 37 ° C với 5% CO 2 .
Thuốc thử và thử nghiệm khả năng sống của tế bào
CBD (Sigma Aldrich, St. Louis, MO, USA; C7515) được cung cấp bởi Giáo sư Tiến sĩ Sang Cheul Oh từ bệnh viện Đại học Guro Hàn Quốc, Hàn Quốc. Bột CBD được hòa tan trong ethanol tuyệt đối (EtOH) và sau đó được bảo quản ở – 20 ° C cho đến khi sử dụng. Đối với các thử nghiệm khả năng sống của tế bào, tế bào HNSCC và HOK được gieo hạt (1 × 10 4 tế bào / giếng) trong đĩa 96 giếng, và sau đó được nuôi cấy trong môi trường tăng trưởng trong 24 giờ. Các tế bào được tiếp xúc với CBD trong môi trường không có huyết thanh trong 24, 48 hoặc 72 giờ trong các điều kiện được chỉ định và sau đó khả năng sống của tế bào được đo bằng Bộ đếm tế bào 8 (CCK-8) (Phòng thí nghiệm Dojindo, Kumamoto, Nhật Bản), theo giao thức của nhà sản xuất, như được mô tả 39. Thuốc chống ung thư bao gồm Cisplatin (C2210000), 5-Fluorouracil (5-FU) (F6627) và Paclitaxel (Taxol) (T1912) đã được mua từ Sigma và được hòa tan trong 0,9% natri clorua, EtOH và DMSO, tương ứng. Chloroquine (CQ) cũng được lấy từ Sigma Aldrich (C6628), được hòa tan trong EtOH, và được bảo quản ở nhiệt độ phòng cho đến khi sử dụng.
Phân tích Isobologram được thực hiện như đã mô tả trước đây 40 . Các giá trị Chỉ số kết hợp (CI) được tính toán dựa trên sự tương tác của hai loại thuốc đã chọn ở nửa nồng độ ức chế (IC 50 ) và sau đó chỉ ra tính đối kháng (CI = 1), tính cộng thêm (CI> 1), hoặc sức mạnh tổng hợp (CI <1) ) về phân tích Isobologram được trình bày bằng đồ thị. Phần mềm ED50 plus v 1.0 được sử dụng để xác định IC 50 của từng loại thuốc như đã mô tả 41 trước đây .
Các xét nghiệm di cư và xâm nhập tế bào
Đối với các thử nghiệm di cư và xâm nhập tế bào, các tế bào xuyên thủng 6,5 mm có chèn màng polycarbonate lỗ 8,0 µm (BD Biosciences, Franklin Lakes, New Jersey, USA; 3403) đã được sử dụng trong nghiên cứu này, theo quy trình như mô tả 42 . Tóm lại, các tế bào được xử lý trước với CBD trong 24 giờ như được chỉ ra ở các nồng độ khác nhau và sau đó là 1,5 × 10 4tế bào / giếng trong 100 µL môi trường DMEM không chứa huyết thanh (Lonza) được thêm vào trong buồng trên. Đối với các thử nghiệm xâm lấn, 10 µg / mL Matrigel (Gibco) được thêm vào buồng trên và được polyme hóa ở 37 ° C trong 2 giờ, và sau đó lượng tế bào được chỉ định được tạo hạt. Buồng dưới của giếng truyền được chứa đầy 600 µL môi trường chứa 20% FBS. Các tế bào được phép di chuyển hoặc xâm nhập trong 24 giờ, và sau đó các tế bào di chuyển được nhuộm bằng nhuộm Diff-Quick (Sysmex, Kobe, Nhật Bản; 24606), theo quy trình như mô tả 43 , và quan sát dưới kính hiển vi.
Thử nghiệm nhuộm màu xanh trypan
Tế bào FaDu (2 × 10 4 tế bào / giếng) được cấy vào các đĩa 24 giếng và được duy trì trong môi trường hoàn chỉnh trong 24 giờ. Ngày hôm sau, các tế bào được xử lý với 0, 0,25, 2,5, 10 hoặc 20 µM CBD trong 24 giờ. Dung dịch xanh trypan đã chuẩn bị (0,4%) (Sigma; T8154) được sử dụng để phân biệt giữa các tế bào sống được và không sống được.
Mô hình khối u xenograft chuột
Các nghiên cứu trên động vật đã được Ủy ban chăm sóc và sử dụng động vật của tổ chức (IACUC) phê duyệt và tất cả động vật được duy trì trong cơ sở động vật Không có mầm bệnh cụ thể (SPF) theo hướng dẫn của IACUC Hàn Quốc. Những con chuột cái 6 tuần tuổi Thiếu Miễn dịch (BALB / c) được mua từ Orient Bio (Seoung-Nam, Hàn Quốc) và được sử dụng trong nghiên cứu in vivo này. Năm con chuột được thử nghiệm trong mỗi nhóm. FaDu (2 × 10 6) tế bào được tiêm dưới da hoặc vào lưỡi và chuột sau đó được nuôi trong 3 ngày. Đối với mô hình khối u được cấy ghép dưới da, CBD được dùng bằng đường uống với liều lượng 5 mg / kg thể trọng trong 4 lần trong một tuần. Những con chuột được cấy tế bào FaDu vào lưỡi của chúng được bổ sung vào CBD (5 mg / kg) bằng cách tiêm trong phúc mạc (ip) 3 lần mỗi tuần. Cisplatin (2,5 mg / kg) được tiêm trong màng bụng hàng tuần. Kích thước khối u đã được xác định mỗi 2-3 ngày và tính theo công thức: V = L × W 2 /2 như mô tả 44 . Các khối u được cân sau khi con vật được hiến tế lúc 4 tuần.
Thử nghiệm hình thành khuẩn lạc
Tế bào FaDu (1 × 10 2 ) được cấy vào đĩa 6 giếng, sau đó được xử lý bằng CBD hoặc thuốc chống ung thư. Sau 2 tuần, các khuẩn lạc được cố định trong 4% paraformaldehyde trong 5 phút và nhuộm bằng thuốc tím pha lê (Sigma; V5265). Các khuẩn lạc được tính toán và chuẩn hóa thành số khuẩn lạc trong nhóm đối chứng không được xử lý.
Xét nghiệm apoptosis
Xét nghiệm apoptosis cho HNSCC được thực hiện bằng cách sử dụng bộ phát hiện FITC Annexin V / Propidium Iodide (PI) (BD Biosciences) như mô tả 42 , và được xác định bằng phương pháp đo tế bào dòng chảy. Sau khi xử lý với CBD 0, 3, 6 hoặc 10 µM trong 24 giờ, tế bào được thu hoạch và trộn với Annexin V và PI trong bóng tối trong 30 phút ở nhiệt độ phòng.
Tách chiết RNA và định lượng thời gian thực RT-PCR
RNA được thu nhận từ HNSCC bằng cách sử dụng thuốc thử TRIZOL (Invitrogen, Carlsbad, CA, USA) để phân tích gen biểu hiện khác biệt. cDNA được phiên mã ngược sử dụng bộ tổng hợp cDNA sợi 1 PrimeScrip (Takara Bio, Tokyo, Nhật Bản) từ 1 μg RNA, theo hướng dẫn của nhà sản xuất. Định lượng phản ứng chuỗi polymerase thời gian thực (qRT-PCR) được thực hiện trong Hệ thống phát hiện ABI Prism 7300 (Hệ thống sinh học ứng dụng) sử dụng thuốc nhuộm xanh ABI SYBR (Hệ thống sinh học ứng dụng, Thành phố Foster, CA, Hoa Kỳ). Phương pháp 2 (−∆∆Ct) được sử dụng để phân tích mức độ biểu hiện tương đối của mRNA và các giá trị được chuẩn hóa theo GAPDH 45 . Các trình tự mồi cụ thể được thể hiện trong quy trình thí nghiệm bổ sung.
Phân tích Western blot
Tế bào HNSCC được nuôi cấy trong đĩa nuôi cấy 6 giếng với kích thước 5 × 10 5tế bào / giếng và tiếp xúc với CBD trong môi trường không có FBS và được ủ thêm trong 24 giờ. Các tế bào thu thập được ly giải và sau đó một lượng protein tương tự được nạp bằng điện di trên gel polyacrylamide natri dodecyl sulfat 8–12%. Các protein được chuyển sang màng, sau đó được chặn bằng sữa tách béo 5% (w / v) chứa TBST (494 mM Tris, 2,74 M NaCl, 54 mM KCl, được điều chỉnh đến pH 7,4 bằng cách sử dụng HCl) trong 30 phút ở nhiệt độ phòng. Các đốm màng được khảo sát bằng các kháng thể chính chống lại PARP bị phân cắt (1: 1000; Công nghệ tín hiệu tế bào, Danvers, Massachusetts, Hoa Kỳ; 5625S), phân cắt Caspase-7 (1: 1000; Công nghệ tín hiệu tế bào; 8438S), LC3 (1: 1000 ; Công nghệ tín hiệu tế bào; 4108S), và β-actin (1: 2000; Công nghệ sinh học Santa Cruz, Dallas, Texas, Hoa Kỳ; sc-47778). Ngày hôm sau, các màng được ủ với kháng thể thứ cấp chống lại IgG liên hợp với peroxidase của cây cải ngựa (Invitrogen, 1: 5000) trong 1 giờ ở nhiệt độ phòng và sau đó hình ảnh được chụp bằng Hệ thống Hình ảnh Fusion Solo (Vilber Lourmat, Marne -La-Vallée, Pháp). Gel và blots có độ dài đầy đủ được đưa vào quy trình thử nghiệm bổ sung.
RNA-seq
RNA tổng số được phân lập từ các dòng tế bào FaDu, Hep2 và SCC15 hoặc chưa được xử lý hoặc được xử lý bằng CBD sử dụng thuốc thử TRIZOL (Invitrogen) và sau đó được định lượng trong máy quang phổ Nanodrop ND-2000. Độ tinh khiết của RNA được kiểm tra bằng Máy phân tích sinh học Agilent 2100. Thư viện giải trình tự Ion Torrent đã được chuẩn bị, theo giao thức bộ chuẩn bị Thư viện AmpliSeq (Thermo Fisher, Phần # MAN0010742) như được mô tả 46. An Ion AmpliSeqTM Transcriptome library was constructed with the Ion Transcriptome Human Gene Expression Kit (Thermo Fisher, SKU # A26325) as per manufacturer’s protocol. 50 ng of total RNA was used for cDNA library preparation and amplified by the Ion AmpliSeq Transcriptome Human Gene Expression kit (over 20,000 human RefSeq genes). Adapters and barcodes were ligated to the library amplicons followed by magnetic bead purification. The concentrated library was measured using an Ion Library TaqMan Quantitation Kit (Thermo Fisher, SKU#4468802), according to the manufacturer’s procedure. Multiple libraries were combined together with equal molar ratios for one Ion 550 chip and clonally amplified using the Ion Chef System (Thermo Fisher), and then sequenced on an Ion Torrent S5xI machine (Thermo Fisher).
Trình tự trong các vùng mục tiêu của một số bản sao Ampliseq được đọc bằng Chương trình căn chỉnh ánh xạ Torrent (TMAP) và trình tự được kiểm soát chất lượng bởi Casava v1.8.2 Pipeline và Cutadapt v1.6 47 , 48 . Các gen kết quả đã được bình thường hóa và kiểm tra sự biểu hiện đặc trưng bằng cách sử dụng DESeq2 và gói Bioconductor 49 , 50 . Hiệu chỉnh Benjamini và Hochberg được sử dụng để thu được giá trị p đã điều chỉnh <0,05. Các gen được điều chỉnh lên và xuống theo gấp đôi hoặc nhiều hơn, mỗi gen được đánh giá để phân tích chức năng và làm giàu lộ trình, chẳng hạn như Bản thể học gen (GO) bằng cách sử dụng tài nguyên tin sinh học Cơ sở dữ liệu cho chú thích, hình ảnh hóa và khám phá tích hợp (DAVID) 51. Bản đồ nhiệt được tạo bằng Multi Experiment Viewer 4.9.0 (MeV 4.9.0) 52 .
Phân tích thống kê
Tất cả dữ liệu đại diện được trình bày dưới dạng trung bình ± độ lệch chuẩn (SD) từ ít nhất trong các thí nghiệm ba lần. Phân tích thống kê được thực hiện bằng cách sử dụng bài kiểm tra t hai phía của học sinh và ANOVA một chiều trong phần mềm SPSS. Giá trị p <0,05 (*), 0,01 (**) và 0,001 (***) được coi là có ý nghĩa thống kê.
Người giới thiệu
- 1.Shrivastava, S. và cộng sự. Xác định dấu hiệu phân tử của các tế bào giống ung thư đầu và cổ. Khoa học. Rep. 5 , 7819. https://doi.org/10.1038/srep07819 (2015).
CAS Bài báo PubMed PubMed Central Google Scholar
- 2.Vigneswaran, N. & Williams, MD Xu hướng dịch tễ học trong ung thư đầu cổ và hỗ trợ chẩn đoán. Răng hàm mặt. Phẫu thuật. Clin. N. Am. 26 , 123–141. https://doi.org/10.1016/j.coms.2014.01.001 (2014).
Bài báo Google Scholar
- 3.Srivastava, G. et al. Hoạt động chống ung thư của kẽm pyrithione trong tế bào ung thư miệng được xác định trong màn hình phân tử nhỏ và mô hình xenograft: ý nghĩa đối với liệu pháp điều trị ung thư miệng. Mol Oncol. 9 , 1720–1735. https://doi.org/10.1016/j.molonc.2015.05.005 (2015).
CAS Bài báo PubMed PubMed Central Google Scholar
- 4.Khan, Z. và cộng sự. Ức chế tăng trưởng và gây mẫn cảm với phóng xạ hóa trị của ung thư biểu mô tế bào vảy ở đầu và cổ (HNSCC) bởi vi-rút sót-siRNA. Radiother. Oncol. https://doi.org/10.1016/j.radonc.2015.12.007 (2015).
Bài báo PubMed Google Scholar
- 5.Dasari, S. & Tchounwou, PB Cisplatin trong điều trị ung thư: cơ chế hoạt động của phân tử. Eur. J. Pharmacol. 740 , 364–378. https://doi.org/10.1016/j.ejphar.2014.07.025 (2014).
CAS Bài báo PubMed Google Scholar
- 6.Feldman, R. và cộng sự. Cấu hình phân tử của ung thư biểu mô tế bào vảy ở đầu và cổ. Cổ đầu 38 (Bổ sung 1), E1625 – E1638. https://doi.org/10.1002/hed.24290 (2016).
Bài báo PubMed Google Scholar
- 7.Larkins, E. và cộng sự. Tóm tắt phê duyệt của FDA: pembrolizumab để điều trị ung thư biểu mô tế bào vảy ở đầu và cổ tái phát hoặc di căn với sự tiến triển của bệnh trong hoặc sau hóa trị liệu có chứa bạch kim. Bác sĩ ung thư 22 , 873–878. https://doi.org/10.1634/theoncologist.2016-0496 (2017).
CAS Bài báo PubMed PubMed Central Google Scholar
- số 8.Dầu Shannon, S. & Opila-Lehman, J. Cannabidiol để giảm sử dụng cần sa gây nghiện: một báo cáo trường hợp. Integr. Med. (Encinitas, CA) 14 , 31–35 (2015).
Google Scholar
- 9.Prud’homme, M., Cata, R. & Jutras-Aswad, D. Cannabidiol như một biện pháp can thiệp cho các hành vi gây nghiện: xem xét có hệ thống các bằng chứng. Chất nền. Lạm dụng 9 , 33–38. https://doi.org/10.4137/SART.S25081 (2015).
CAS Bài báo PubMed PubMed Central Google Scholar
- 10.Bruni, N. và cộng sự. Hệ thống phân phối cannabinoid để điều trị đau và viêm. Phân tử 23 , 2478. https://doi.org/10.3390/molecules23102478 (2018).
CAS Bài báo PubMed Central Google Scholar
- 11.Hayakawa, K., Mishima, K. & Fujiwara, M. Tiềm năng điều trị của cannabidiol không hướng thần trong đột quỵ do thiếu máu cục bộ. Dược phẩm (Basel, Thụy Sĩ) 3 , 2197–2212. https://doi.org/10.3390/ph3072197 (2010).
CAS Bài báo Google Scholar
- 12.Maroon, J. & Bost, J. Đánh giá về lợi ích thần kinh của phytocannabinoids. Phẫu thuật. Neurol. Int. 9 , 91–91. https://doi.org/10.4103/sni.sni_45_18 (2018).
Bài báo PubMed PubMed Central Google Scholar
- 13.Blázquez, C. và cộng sự. Ức chế sự hình thành mạch khối u bởi cannabinoids. FASEB J. 17 , 529–531. https://doi.org/10.1096/fj.02-0795fje (2003).
CAS Bài báo PubMed Google Scholar
- 14.Qamri, Z. và cộng sự. Chất chủ vận thụ thể cannabinoid tổng hợp ức chế sự phát triển của khối u và sự di căn của ung thư vú. Mol Bệnh ung thư. 8 , 3117–3129. https://doi.org/10.1158/1535-7163.MCT-09-0448 (2009).
CAS Bài báo PubMed PubMed Central Google Scholar
- 15.Śledziński, P., Zeyland, J., Słomski, R. & Nowak, A. Tình trạng hiện tại và viễn cảnh tương lai của cannabinoids trong sinh học ung thư. Thuốc điều trị ung thư. 7 , 765–775. https://doi.org/10.1002/cam4.1312 (2018).
Bài báo PubMed PubMed Central Google Scholar
- 16.Russo, EB Cannabinoids trong quản lý cơn đau khó điều trị. Họ. Clin. Quản lý rủi ro. 4 , 245–259. https://doi.org/10.2147/tcrm.s1928 (2008).
CAS Bài báo PubMed PubMed Central Google Scholar
- 17.Liu, C. và cộng sự. Cannabinoids thúc đẩy sự tiến triển của ung thư biểu mô tế bào vảy ở đầu và cổ dương tính với HPV thông qua kích hoạt p38 MAPK. J. Clin. Ung thư Res. 26 , 2693–2703. https://doi.org/10.1158/1078-0432.CCR-18-3301 (2020).
CAS Bài báo Google Scholar
- 18.Lai, P., Chi, T.-Y. & Chen, G. Các mức độ khác nhau của p53 gây ra quá trình apoptosis hoặc bắt giữ chu kỳ tế bào trong một dòng tế bào ung thư biểu mô tế bào gan điều hòa doxycycline trong ống nghiệm. Apoptosis 12 , 387–393. https://doi.org/10.1007/s10495-006-0571-1 (2007).
CAS Bài báo PubMed Google Scholar
- 19.Liu, J. và cộng sự. Điều hòa giảm của GADD45A tăng cường độ nhạy với hóa chất trong u ác tính. Khoa học. Đại diện https://doi.org/10.1038/s41598-018-22484-6 (2018).
Bài báo PubMed PubMed Central Google Scholar
- 20.Stewart-Ornstein, J. & Lahav, G. Động lực học của CDKN1A trong các tế bào đơn lẻ được xác định bởi bộ công cụ gắn thẻ huỳnh quang nội sinh. Ô. Bản tái bản 14 , 1800–1811. https://doi.org/10.1016/j.celrep.2016.01.045 (2016).
CAS Bài báo PubMed PubMed Central Google Scholar
- 21.Valastyan, S. & Weinberg, RA Di căn khối u: hiểu biết về phân tử và các mô hình phát triển. Tế bào 147 , 275-292. https://doi.org/10.1016/j.cell.2011.09.024 (2011).
CAS Bài báo PubMed PubMed Central Google Scholar
- 22.Lu, DY và cộng sự. Phối hợp thuốc trong điều trị ung thư trên lâm sàng. Rev. Clin gần đây. Thử nghiệm 12 , 202–211. https://doi.org/10.2174/1574887112666170803145955 (2017).
CAS Bài báo PubMed Google Scholar
- 23.Shrivastava, A., Kuzontkoski, PM, Groopman, JE & Prasad, A. Cannabidiol gây chết tế bào theo chương trình trong các tế bào ung thư vú bằng cách phối hợp trao đổi chéo giữa apoptosis và autophagy. Mol Bệnh ung thư. 10 , 1161–1172. https://doi.org/10.1158/1535-7163.Mct-10-1100 (2011).
CAS Bài báo PubMed Google Scholar
- 24.Jeong, S. và cộng sự. Quá trình chết rụng do cannabidiol được thực hiện qua trung gian kích hoạt Noxa trong các tế bào ung thư đại trực tràng ở người. Chữ cái ung thư. 447 , 12–23. https://doi.org/10.1016/j.canlet.2019.01.011 (2019).
CAS Bài báo PubMed Google Scholar
- 25.McKallip, RJ và cộng sự. Quá trình apoptosis do cannabidiol gây ra ở các tế bào bệnh bạch cầu ở người: một vai trò mới của cannabidiol trong việc điều hòa biểu hiện p22phox và Nox4. Mol Pharmacol. 70 , 897–908. https://doi.org/10.1124/mol.106.023937 (2006).
CAS Bài báo PubMed Google Scholar
- 26.Carracedo, A. và cộng sự. Cannabinoid gây ra quá trình apoptosis của các tế bào khối u tuyến tụy thông qua các gen liên quan đến căng thẳng mạng lưới nội chất. Ung thư Res. 66 , 6748–6755. https://doi.org/10.1158/0008-5472.Can-06-0169 (2006).
CAS Bài báo PubMed Google Scholar
- 27.Salazar, M. và cộng sự. Hoạt động cannabinoid gây chết tế bào qua trung gian autophagy thông qua kích thích căng thẳng ER trong tế bào u thần kinh đệm ở người. J. Clin. Đầu tư. 119 , 1359–1372. https://doi.org/10.1172/jci37948 (2009).
CAS Bài báo PubMed PubMed Central Google Scholar
- 28.Armstrong, JL và cộng sự. Khai thác quá trình tự thực hiện gây độc tế bào do cannabinoid gây ra để thúc đẩy tế bào u ác tính chết. J. Đầu tư. Dermatol. 135 , 1629–1637. https://doi.org/10.1038/jid.2015.45 (2015).
CAS Bài báo PubMed Google Scholar
- 29.Erener, S. và cộng sự. Caspase 7 được kích hoạt bởi ảnh hưởng phân cắt PARP1 để tăng cường sự biểu hiện của một tập hợp con các gen mục tiêu NF-κB. Mol Tế bào 46 , 200-211. https://doi.org/10.1016/j.molcel.2012.02.016 (2012).
CAS Bài báo PubMed Google Scholar
- 30.Kidger, AM & Keyse, SM Cơ chế điều hòa tín hiệu Ras / ERK gây ung thư bằng cách kích hoạt mitogen phosphatases protein kinase (MKPs) có tính đặc hiệu kép. Semin. Ô. Nhà phát triển. Biol. 50 , 125–132. https://doi.org/10.1016/j.semcdb.2016.01.009 (2016).
CAS Bài báo PubMed PubMed Central Google Scholar
- 31.Sebolt-Leopold, JS & Herrera, R. Nhắm mục tiêu theo dòng protein kinase kích hoạt mitogen để điều trị ung thư. Nat. Rev. Cancer 4 , 937–947. https://doi.org/10.1038/nrc1503 (2004).
CAS Bài báo PubMed Google Scholar
- 32.Shen, J. và cộng sự. DUSP1 ức chế tăng sinh tế bào, di căn và xâm lấn và hình thành mạch trong ung thư túi mật. Mục tiêu 8 , 12133–12144. https://doi.org/10.18632/oncotarget.14815 (2017).
Bài báo PubMed PubMed Central Google Scholar
- 33.Wang, J., Zhou, J.-Y., Kho, D., Reiners, JJ Jr. & Wu, GS Vai trò đối với DUSP1 (protein phosphatase đặc hiệu kép 1) trong cơ chế điều hòa autophagy. Autophagy 12 , 1791–1803. https://doi.org/10.1080/15548627.2016.1203483 (2016).
CAS Bài báo PubMed PubMed Central Google Scholar
- 34.Hsu, L.-S. et al. KLF6 ức chế sự di cư và xâm lấn của ung thư miệng thông qua việc điều chỉnh giảm các dấu ấn trung mô và ức chế các hoạt động của MMP-9. Int. J. Med. Khoa học. 14 , 530–535. https://doi.org/10.7150/ijms.19024 (2017).
CAS Bài báo PubMed PubMed Central Google Scholar
- 35.Beck, TN và cộng sự. EGFR và RB1 làm dấu ấn sinh học kép trong ung thư đầu cổ âm tính với HPV. Mol Bệnh ung thư. 15 , 2486–2497. https://doi.org/10.1158/1535-7163.Mct-16-0243 (2016).
CAS Bài báo PubMed PubMed Central Google Scholar
- 36.Sangodkar, J. và cộng sự. Nhắm mục tiêu trục FOXO1 / KLF6 điều chỉnh phản ứng điều trị và tín hiệu EGFR. J. Clin. Đầu tư. 122 , 2637–2651. https://doi.org/10.1172/JCI62058 (2012).
CAS Bài báo PubMed PubMed Central Google Scholar
- 37.Hellingay, T. Cơ chế cơ bản của khả năng sát thương tổng hợp PARP và BRCA: giải tỏa những hiểu lầm. Mol Oncol. 5 , 387–393. https://doi.org/10.1016/j.molonc.2011.07.001 (2011).
CAS Bài báo PubMed PubMed Central Google Scholar
- 38Cheung, CHY và cộng sự. Mạng lưới chức năng điều chỉnh MCM2 trong ung thư phổi bằng phương pháp tiếp cận protein đa chiều. Khoa học. Bản 7 , 13302–13302. https://doi.org/10.1038/s41598-017-13440-x (2017).
QUẢNG CÁO CAS Bài báo PubMed PubMed Central Google Scholar
- 39.Takanezawa, Y., Nakamura, R., Sone, Y., Uraguchi, S. & Kiyono, M. Chế độ tự thực phụ thuộc Atg5 đóng vai trò bảo vệ chống lại độc tế bào do methylmercury gây ra. Toxicol. Lett. 262 , 135–141. https://doi.org/10.1016/j.toxlet.2016.09.007 (2016).
CAS Bài báo PubMed Google Scholar
- 40.Chou, TC Các nghiên cứu kết hợp thuốc và định lượng sức mạnh tổng hợp của chúng bằng phương pháp Chou – Talalay. Ung thư Res. 70 , 440–446. https://doi.org/10.1158/0008-5472.Can-09-1947 (2010).
CAS Bài báo PubMed Google Scholar
- 41.Offman, MN và cộng sự. Kỹ thuật hợp lý của L-asparaginase cho thấy tầm quan trọng của hoạt tính kép đối với độc tính của tế bào ung thư. Máu 117 , 1614–1621. https://doi.org/10.1182/blood-2010-07-298422 (2011).
CAS Bài báo PubMed Google Scholar
- 42.Hu, A. và cộng sự. Curcumin như một liệu pháp điều trị ung thư biểu mô tế bào vảy ở đầu và cổ bằng cách kích hoạt SIRT1. Khoa học. Rep. 5 , 13429. https://doi.org/10.1038/srep13429 (2015).
QUẢNG CÁO CAS Bài báo PubMed PubMed Central Google Scholar
- 43.Nishimura, Y. và cộng sự. Sự biểu hiện quá mức của YWHAZ liên quan đến sự tăng sinh tế bào khối u và kết quả ác tính của ung thư biểu mô dạ dày. Br. J. Ung thư 108 , 1324–1331. https://doi.org/10.1038/bjc.2013.65 (2013).
CAS Bài báo PubMed PubMed Central Google Scholar
- 44.Faustino-Rocha, A. và cộng sự. Ước tính thể tích khối u tuyến vú của chuột bằng cách sử dụng thước cặp và đo siêu âm. Lab Anim. 42 , 217–224. https://doi.org/10.1038/laban.254 (2013).
Bài báo Google Scholar
- 45.Livak, KJ & Schmittgen, TD Phân tích dữ liệu biểu hiện gen tương đối bằng cách sử dụng PCR định lượng thời gian thực và Phương pháp 2 (-Delta Delta C (T)). Phương pháp (San Diego, CA) 25 , 402–408. https://doi.org/10.1006/meth.2001.1262 (2001).
CAS Bài báo Google Scholar
- 46.Dyjack, N. và cộng sự. Dải băng da xâm lấn tối thiểu Giải trình tự RNA xác định các đặc điểm mới của bệnh viêm da dị ứng cấp độ 2 type 2. J. Clin dị ứng. Immunol. 141 , 1298–1309. https://doi.org/10.1016/j.jaci.2017.10.046 (2018).
CAS Bài báo PubMed PubMed Central Google Scholar
- 47.Martin, M. CUTADAPT loại bỏ trình tự bộ điều hợp khỏi các lần đọc trình tự thông lượng cao. EMBnet. J. https://doi.org/10.14806/ej.17.1.200 (2011).
Bài báo Google Scholar
- 48.Nakazato, T., Ohta, T. & Bono, H. Khai thác chức năng dựa trên thiết kế thử nghiệm và mô tả đặc điểm của dữ liệu giải trình tự thông lượng cao trong kho lưu trữ đọc trình tự. PLoS ONE 8 , e77910. https://doi.org/10.1371/journal.pone.0077910 (2013).
QUẢNG CÁO CAS Bài báo PubMed PubMed Central Google Scholar
- 49.Love, M., Huber, W. & Anders, S. Ước tính vừa phải sự thay đổi nếp gấp và sự phân tán đối với dữ liệu RNA-Seq với DESeq2. Genome Biol 15 , 550. https://doi.org/10.1186/s13059-014-0550-8 (2014).
CAS Bài báo PubMed PubMed Central Google Scholar
- 50.Huber, W. và cộng sự. Điều phối phân tích bộ gen thông lượng cao với chất dẫn sinh học. Nat. Phương pháp 12 , 115–121. https://doi.org/10.1038/nmeth.3252 (2015).
CAS Bài báo PubMed PubMed Central Google Scholar
- 51.Huang, DW và cộng sự. Tài nguyên tin sinh học DAVID: cơ sở dữ liệu chú thích mở rộng và các thuật toán mới để trích xuất sinh học tốt hơn từ danh sách gen lớn. Nucl. Axit Res. 35 , W169 – W175. https://doi.org/10.1093/nar/gkm415 (2007).
Bài báo PubMed Google Scholar
- 52.Phân tích Howe, EA, Sinha, R., Schlauch, D. & Quackenbush, J. RNA-Seq trong MeV. Tin sinh học 27 , 3209–3210. https://doi.org/10.1093/bioinformatics/btr490 (2011).
CAS Bài báo PubMed PubMed Central Google Scholar
Tải xuống tài liệu tham khảo
Sự nhìn nhận
Công trình này được hỗ trợ bởi khoản tài trợ của Đại học Hàn Quốc và khoản tài trợ của Quỹ Nghiên cứu Quốc gia Hàn Quốc (NRF) do chính phủ Hàn Quốc (MSIT) tài trợ (Số 2017R1A2B4005163).
Thông tin tác giả
Chi nhánh
- Khoa Tai Mũi Họng-Phẫu thuật Đầu và Cổ, Bệnh viện Đại học Hàn Quốc Guro, 80 Guro-dong, Guro-gu, Seoul, 08308, Hàn Quốc
Yoon Young Go, Su Ra Kim, Do Yeon Kim, Sung-Won Chae & Jae-Jun Song
- Viện Trung tâm Hội tụ Chăm sóc Sức khỏe, Bệnh viện Đại học Hàn Quốc Guro, Seoul, Hàn Quốc
Yoon Young Go & Jae-Jun Song