Phương pháp điều trị ung thư của bác sĩ Hoàng Xuân Ba gây tranh cãi

Chứng kiến sự bất lực của một người con với nỗi đau từ căn bệnh nan y của bố, bác sỹ Hoàng Xuân Ba đã đưa ra một quyết định táo bạo với y học lúc đó, để sau này ông trở thành chiến thắng tử thần giành giật sự sống cho nhiều bệnh nhân mắc bệnh ung thư.
Chứng kiến sự bất lực của một người con với nỗi đau từ căn bệnh nan y của bố, bác sỹ Hoàng Xuân Ba đã đưa ra một quyết định táo bạo với y học lúc đó, để sau này ông trở thành chiến thắng tử thần giành giật sự sống cho nhiều bệnh nhân mắc bệnh ung thư.
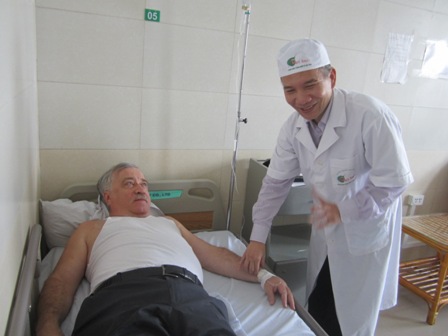 |
| Bác sỹ Hoàng Xuân Ba khám cho một bệnh nhân ngoại quốc |
Những nỗ lực vượt thời gian và thành công không tưởng…
Từ năm 1995, Bác sỹ Hoàng Xuân Ba đã khá nổi tiếng ở Viện Lê – nin – grat với những thành công trong một số công trình nghiên cứu khoa học, trong đó có công trình nghiên cứu cách điều trị bệnh hen phế quản.
Tình cờ, một đồng nghiệp người Nga nhờ anh điều trị giảm đau cho bố cô, vốn bị ung thư giai đoạn cuối đang bị tắc nghẽn và tràn dịch màng phổi. Sau khi khám cho bệnh nhân, bác sỹ Ba đã quyết định đưa ra một phác đồ điều trị với mục đích để bệnh nhân đỡ đau và dễ thở hơn.
Thật bất ngờ, bệnh nhân đã hồi phục sức khỏe một cách rất nhanh chóng. 6 tháng sau khám lại, khối u của BN chỉ còn chưa đầy 10%, hiện tượng tràn dịch màng phổi cũng không còn nữa..
Với mong muốn chinh phục những căn bệnh nan y, Hoàng Xuân Ba đã dồn hết tâm sức và tiền của kiếm được vào việc nghiên cứu công trình khoa học này. Trời đã không phụ lòng người, công sức, thời gian và nỗ lực của anh đã được đền đáp xứng đáng.
Năm 1996, BS Ba được mời tham gia một hội thảo tại Mỹ và báo cáo tham luận về xuất huyết giảm tiểu cầu. Cơ chế khoa học của đề này đối lập hoàn toàn với quan điểm và cách thức điều trị bệnh của các nhà chuyên môn thời bấy giờ, nhưng bằng những lập luận sắc bén và kiến thức y học chắc chắn cộng với những thành công rút ra trong quá trình thực nghiệm, Ba đã thuyết phục được Hội đồng khoa học.
Sau thành công tại hội nghị này, Xuân Ba được một hãng nghiên cứu công nghệ kỹ thuật miễn dịch của Mỹ mời về làm việc và được nhập cư vào Mỹ theo diện Visa-01 (dành cho các cá nhân có khả năng đặc biệt trong khoa học).
Và cũng chính trong thời gian làm việc tại đây, anh đã tập trung mọi tâm sức vào nghiên cứu hướng điều trị mới cho các bênh nhân ung thư. Các kết quả khả quan trong phương pháp nghiên cứu khoa học này lại một lần nữa gây chấn động dư luận cũng như giới khoa học thế giới.
Đích thân GS. Fritz Sorgel, đồng chủ trì Hội nghị quốc tế lần thứ II về ung thư đã có lời mời TS. BS. Hoàng Xuân Ba sang báo cáo tại Nurenberg (Đức) về giả thuyết nguyên nhân gây ung thư và cách điều trị.
Nói về nguyên tắc của việc điều trị căn bệnh nan y này, BS Ba chia sẻ: “Nguyên tắc của tôi là tránh xa tàn sát, tiêu diệt tế bào ung thư như đang làm hiện nay, mà cung cấp nguyên liệu giúp tạo năng lượng đầy đủ cho tế bào ung thư và chống lại sự kích thích theo cơ chế thần kinh ở màng tế bào và u tạo thành các gốc tự do mới sinh…”.
“Suốt đời hành nghề trong sự vô tư và thân thiết…
Phương pháp giảm đau và chữa triệu chứng đối với bệnh nhân ung thư của TS. BS. Hoàng Xuân Ba đã đạt được nhưng thành công bước đầu, qua thực tế điều trị cho bệnh nhân tại Mỹ.
Với mục đích chuyển giao kỹ thuật mới này và hỗ trợ đồng bào Việt Nam chống chọi và tiến tới chiến thắng căn bệnh nan y này, anh đã bỏ qua rất nhiều lời mời cộng tác hấp dẫn để về nước giúp đỡ người bệnh nan y nghèo. Những nơi mà anh từng hợp tác là Trung tâm Y tế Lao động (Bộ NN & PTNT), Khoa U bướu (Viện 198, Bộ Công an) và hiện anh đang làm việc tại Trung tâm Y khoa Quốc tế Việt Xuân, Viện Y học cổ truyền, Bộ Công an.
Và đã có hàng ngàn bệnh nhân nan y được hưởng lợi từ phương pháp này. Thậm chí, có đến hàng trăm bệnh nhân là người nước ngoài nghe danh anh đã tìm đến để được anh khám, chữa bệnh và đã giành giật được sự sống từ tay tử thần.
Hơn hai năm cộng tác với Trung tâm Y tế Lao động và Khoa U bướu, Viện 198 bác sỹ Hoàng Xuân Bua không hề nhận một đồng lương, mà chỉ muốn chuyển giao cho các cơ sở phương pháp khoa học của mình.
Tại Trung tâm Y khoa Quốc tế Việt Xuân cũng vậy, lúc nào anh cũng hết lòng, tận tụy với người bệnh, với một lời thề sắc son của người chiến sỹ áo trắng: “Tôi suốt đời hành nghề trong sự vô tư và thân thiết”.
Hầu hết các trường hợp khám và điều trị ở đây đều qua tay bác sỹ Ba. Cũng tự tay anh pha chế và điều chỉnh thuốc, chế độ ăn cho từng ca bệnh.
Bệnh nhân nào khó khăn quá anh miễn giảm viện phí, tiền thuốc, thậm chí hỗ trợ kinh phí đi lại, ăn ở những trường hợp quá khó khăn và xa xôi.
Là bác sỹ Tây y nhưng bác sỹ Ba còn tham gia bào chế rất nhiều loại thuốc đông y nổi tiếng thế giới và khu vực, trong đó có Nga phụ khang, Novispes và nhiều loại thuốc tiêm, truyền khác.
Hiện tại, anh đang tập trung, dồn sức tiếp tục nghiên cứu những loại thuốc từ nguồn thảo dược từ thiên nhiên và sản xuất trong nước với mong muốn hạn chế đến mức thấp nhất chi phí từ tiền túi của người dân trong việc khám chữa bệnh.
Kết ngắn
Ra nước ngoài học tập, làm việc ở đó, rồi lại về Việt Nam khám chữa bệnh là một lẽ thường tình. Nhưng, tạo được tiếng vang ở xứ người, rồi lại thu hút được người nước ngoài về Việt Nam trị bệnh lại là một việc không phải ai cũng làm được, chỉ có được ở một người yêu nước, tận tâm, tận lực và có tâm huyết với người bệnh nghèo như BS Hoàng Xuân Ba mà thôi./.
Sau đây là lời giới thiệu của 1 Viện trưởng Viện nghiên cứu dị ứng nói về Bs Ba – người mà bệnh nhân cũng như đồng nghiệp trên thế giới luôn gọi thân thiết là Dr Ba Hoàng
BS. TS Hoàng Xuân Ba – BS trình độ cao nhất về ung thư, miễn dịch, dị ứng, …
Trích giới thiệu của TS. Stephen Levine, giáo sư di truyền học phân tử và viện trưởng viện nghiên cứu dị ứng, California, USA về phương pháp điều trị ung thư của BS Hoàng Xuân Ba tại hội nghị thế giới lần thứ 4 về Y học Dinh dưỡng tại San Francisco.
BS.TS Hoàng Xuân Ba đã được đào tạo tại St.Petersburg với chuyên khoa ung thư, miễn dịch lâm sàng và nhi khoa. Ông ta là một trong những bác sỹ ở trình độ cao nhất mà tôi được biết. Đặc biệt, những đột phá về điều trị của ông trong những năm gần đây ở Mỹ đã trở thành huyền thoại.
BS. Ba đã chữa thành công ung thư phổi, gan, vú và tiền liệt tuyến với kết quả vượt xa những gì được báo cáo trong các tài liệu y khoa. Tôi đã trực tiếp gặp và nói chuyện với một số bệnh nhân để tự khẳng định về kết quả này. Một số bệnh nhân là bạn, cộng tác viên, và người thân trong gia đình tôi. Trong những trường hợp này, tôi đã chứng kiến sự hồi phục của bệnh nhân bị ung thư thận tiền phát , ung thư phổi đã di căn, ung thư tiền liệt tuyến giai đoạn cuối và ung thư vú với di căn vào xương mà các bác sỹ đã khuyên bệnh nhân chỉ dùng thuốc giảm đau cho những ngày cuối cùng.
BS.Ba đã cho tôi biết là ông ta có thể giúp bạn rất thân của tôi bị ung thư thận. Điều này rất không bình thường vì ung thư thận thường được điều trị với hiệu quả rất thấp bằng phẫu thuật và hoá trị liệu. Chỉ trong vòng 45 ngày bạn tôi đã không còn dấu vết của khối u ở thận sau khi làm lại PET scan và cả kiểm chứng chỉ số thực nghiệm về lượng kháng nguyên của carcinoma thận. Một đồng nghiệp của tôi là tiến sỹ về hoá học có người thân bị ung thư phổi đã di căn, chỉ trong vòng 6 tuần đã có được sức khoẻ và thể trạng gần như bình thường và chỉ còn lại một dấu vết nhỏ về khối u trên phim X-quang.
Tỷ lệ thành công của phương pháp chữa bệnh của BS.Ba rất cao. Rõ ràng là ông ta đã tìm thấy những nguyên lý đúng đắn về căn bệnh nan y này và giúp chúng ta có hiểu biết đúng hơn và điều trị nó hiệu quả hơn.Những số liệu và lâm sàng, cận lâm sàng của bệnh nhân sẽ được cung cấp.
Một điều rõ ràng từ kết quả lâm sàng của phương pháp điều trị kể trên là nó có tác dụng chống ung thư mạnh mà không kèm theo một tác dụng độc hại nào. Phương pháp này còn có tác dụng nâng cao thể trạng của bệnh nhân và giảm đau tốt mà không cần đến morphine và các thuốc chống đau khác. Cách chữa bệnh này không làm tổn hại các cơ quan trong cơ thể một chút nào, không gây lệ thuộc và không đắt tiền.
Stephen A. Levine PhD
President, Allergy Research Group
Đây là thông tin về Getwell cùng các sp được sản xuất theo công thức của BS Hoàng Xuân Ba
http://www.getwellnatural.com/about-us.aspx
Dr. Shaw received his medical training at Creighton University of Medicine and is Board Certified in internal medicine. He is a member of the American College for the Advancement in Medicine and has been the director of the Natural Immune Supplements Corporation. Presently, he operates an Integrated Medical Clinic in Los Altos, California and has had great success in blending his knowledge of Eastern and Western modalities. Dr. Shaw specializes in integrated medicine which incorporates the best parts of alternative nutritional therapies using herbs, vitamins, amino acids, and other natural substances and therapies. Dr. Shaw has offered Get Well Natural formulas in his personal practice for many years and with many satisfied patients. In combination with Dr. Ba Hoang, Dr. Graeme Shaw has been published in a number of international medical journals and pivotal publications. *
Thực tế, người VN rất giỏi, rất nhiều nhân tài làm rạng danh tổ quốc. GS Ngô Bảo Châu, và BS Hoàng Xuân Ba cùng ở trong lĩnh vực nghiên cứu KH, 1 ng thì rạng danh về Toán – khoa học cơ bản, 1 ng thì tìm tòi và đưa ra rất nhiều phương thuốc cứu người mà đã được Y học Mỹ – nền y học phát triển nhất thế giới – tôn lên là 1 HUYỀN THOẠI.
Tôi có tập tài liệu ( ngót 40 trang ) ” Hội thảo quốc tế về ung thư lần đầu tiên do BV 19 – 8 (Bộ Công an) tổ chức- Hy vọng mới cho BN ung thư”, trong đó GS Phạm Song, Chủ tịch Hội Y học VN đánh giá
“ Là hội nghị về UT đầu tiên mà tôi thấy thích thú nhất từ năm 2010 đến nay, vì được thấy một “giọng điệu” hoàn toàn khác, khi các nghiên cứu đều rất sâu với những chứng cứ thực tế, sinh động trên bệnh nhân. VN phải đi trên con đường này, may ra mới kịp ”
Tài liệu rất dài (44 trang kể cả đoạn tiếng Anh) , tôi chỉ xin nói một vài điều cơ bản, mà qua đó chúng tôi THEO CHỮA.
1 / Liệu pháp DMSO. Viên đạn kỳ diệu cho bệnh ung thư. (Trích)
Bạn đã bao giờ nghe nói chụp PET Scan chưa ?Khi sử dụng PET Scan kỹ thuật viên sẽ cho bệnh nhân UT một giải pháp phóng xạ glucose (ví dụ: chất phóng xạ để đánh dấu hoặc thêm glucose) Bởi vì các tế bào UT tiêu thụ nhiều glucose hơn các tế bào thường 15 lần, các tế bào UT sẽ hấp thụ phóng xạ glucose hơn các tế bào thường 15 lần. Kết quả là khi họ chụp PET Scan các tế bào UT hiện lên trên tia X quang.
Vì vậy y học chính thống biết làm thế nào để nhằm tới các tế bào UT. Nếu y học hiện nay thật sự quan tâm đến hiệu quả trong việc chữa bệnh UT , bạn có nghĩ rằng họ nên tìm kiếm các cách để tạo glucose theo cách mà glucose nhằm tới các tế bào UT và tiêu diệt chúng ? Nói một cách khác ,bạn có nghĩ rằng ngành y học nên tìm kiếm một cách để nhằm tới các tế bào UT với mục đích tiêu diệt chúng thay vì chỉ để chúng hiện lên trong tia X quang ?
Cũng như cách điều trị UT hiện nay nhưng thay vì sử dụng glucose nó sử dụng DMSO ( Dimethylsul foxide)
1. DMSO gắn với (hóa chất để tấn công) kèm theo các thuốc hóa trị liệu, sau đó
2. DMSO (luôn luôn mục tiêu các tế bào UT) sẽ nhằm mục tiêu , và
3. DMSO sẽ kéo hóa chất vào tế bào UT, và
4. Các hóa chất ( giờ có khả năng nhằm tới các tế bào UT ) sẽ tiêu diệt tế bào UT.
. . . . .
… Với cách điều trị này hóa trị liệu chỉ hướng tới các tế bào UT . Chỉ cần một liều nhỏ hóa trị liệu và không có các tác dụng phụ từ nó bởi vì tất cả các chất hóa trị liệu đều nhằm tới các tế bào UT.
… Bạn có thể hỏi BS chuyên khoa UT của bạn tại sao các cơ hội để sống của bạn chỉ là 3 % (bỏ qua tất cả những thống kê phù phiếm như là tỉ lệ sống là 5 năm và những lời nói dối rằng sẽ giảm bớt và phản ứng) trong khi cơ hội sống sót của bạn có thể lên tới 90 % nếu họ sử dụng DMSO chỉ với một lượng nhỏ hóa chất .
. . . . .
2 / Nhưng không may các BS sử dụng các cách điều trị đơn thuần chỉ dựa trên lợi nhuận thu được hơn là hiệu quả của chúng !
DMSO có độ an toàn rất cao và là sản phẩm 100 % nguồn gốc thiên nhiên từ gỗ. Tất nhiên cũng giống IPT(liệu pháp Insulin) , sự phát hiện ra DMSO đã bị giấu kín vì nó không thể bảo vệ được bản quyền và mang lại lợi nhuận lớn vì có thể sản xuất ra hàng tấn từ công nghiệp chế biến gỗ. Chỉ có tác dụng phụ duy nhất của DMSO là gây ra mùi ở một số bệnh nhân ( mùi ngô non)
Ở HN hiện có một số nơi khám và chữa bệnh theo phương pháp của BS.TS Hoàng Xuân Ba chúng tôi tổng kết được như sau:
1. Trung tâm Y khoa Quốc tế Việt Xuân, 278 Lương Thế Vinh, Hà Nội. Ở đây dịch vụ chuyên nghiệp, thái độ chăm sóc bệnh nhân rất tốt, cơ sở khang trang, hiện đại như khách sạn. Chi phí 300K/giường thường và 800K/giường VIP, không kể thuốc men và công tiêm truyền. Chi phí ở đây hơi cao vì phòng ốc đẹp đẽ và chuyên nghiệp.
2. TT Y tế 178 Thái Hà. Điện thoại BS Tuần – người học trò giỏi của BS Ba 0986234555. Chi phí tiền giường khá thấp, nằm ở Trung tâm thành phố, tiền công tiêm truyền và chăm sóc cũng thấp – chỉ bẳng 1/3 tới 1/4 Việt Xuân, nhưng thái độ phục vụ không được tận tình và chu đáo như Việt Xuân
3. Bệnh viện 19-8 Bộ Công An vào Khoa ung biếu tìm chủ nhiệm khoa : Thượng tá Trần Quốc Hùng ĐT: 0904109756. Tiền phòng và công chăm sóc của y bác sỹ cũng bình thường, tuy nhiên, giống như TT 178 Thái Hà – ỏ đây việc chăm sóc bệnh nhân vẫn còn mang tính nhà nước
4. Việt Medicare 133 Thái Hà, có các bác sỹ
GSTS Lê Đức Hinh, CT Hội TKVN (vào sáng Thứ 2)
GSTS Hà Văn Mạo, CT Hội Tiêu hóa VN ( sáng Thứ 3)
GSTS Trần Đỗ Trinh ,nguyên Viện trưởng Viện Tim, (sáng Thứ 4, đầu tháng)
GSTS Nguyễn Ngọc Tước, nguyên GS ViệnTim TW, (sáng Thứ 4)
BS CK II Nguyễn Thị Cẩm Châu , nguyên P.CN khoa Khớp BV Bạch Mai (sáng Thứ5)
TS Hà Phan Hải An, CN khoa Thận & lọc máu BV Việt Đức;
Tất cả 4 nơi trên đều sử dụng thuốc men và phác đồ điều trị của BS.TS Hoàng Xuân Ba.
Để được khám bệnh, chữa trị bởi BS Ba và các cộng sự, có thể liên lạc theo các địa chỉ sau:
Số điện thoại: 093.432.8998
http://www.dailymoti…-ung-thy_people
ĐƯờng link trên là phóng sự của VTC10 viết về BS Ba e thấy trên dailymotion. BS rất giản dị, rất đời thường, trả lời rất thực, không màu mè – y nhu ngoài đời vậy.
E tìm hiểu trên google thì được biết, kênh truyền hình này có 1 chương trình Kết nối cộng đồng chuyên nói về những người Việt thành danh ở nước ngoài, những hoạt động của họ có tác động và ảnh hưởng nhất định trong cộng đồng người Việt và quốc tế.
E không biết cách chèn đường link vào như các thành viên khác nên copy cả đường dẫn vào ạ, ai biết thì chèn giúp em với, làm thế nào để hiện ra màn hình rồi chỉ việc bấm PLAY xem ấy ạ.
http://www1.vtc.com.vn/view/9/78644/ket_noi_cong_dong__niem_hy_vong_cho_benh_nhan_ung_thu.aspx#/
Đường link này là từ web của VTC
Tương tự với việc đã xảy ra ở Mỹ đầu những năm 1990, ở nước ta trong những tháng vừa qua đã rộ lên một làn sóng gây nghi ngờ và kết tội bừa bãi bất kể sản phẩm phòng và chữa bệnh nào có nguồn gốc thiên nhiên. Đây có phải là những việc làm chỉ vì mục đích bảo vệ người tiêu dùng và sức khỏe của những người dân bình thường không?
Chúng ta cần có một cách nhìn khách quan, tỉnh táo để không lún sâu vào cạm bẫy của những kẻ coi bệnh tật như một mảnh đất màu mỡ để kiếm lời.
Ngay từ năm 1902, Thomas Edison, một nhà sáng chế và phát minh vĩ đại của nhân loại đã nói lên mong ước và cũng là định hướng rất đúng đắn cho y học: “Bác sỹ của tương lai là những người không chỉ cho thuốc mà còn cần chú trọng đến cơ thể của những người bệnh, chế độ dinh dưỡng của họ để tìm ra nguyên nhân bệnh tật và cách phòng ngừa chúng”.
Điều đáng tiếc là với đại bộ phận các bác sỹ hiện nay, họ chỉ biết đến thuốc và thuốc. Càng chỉ định nhiều thuốc mới, thuốc đắt tiền, uy tín và thu nhập của họ càng cao.
Nhiều bệnh nhân bị các bệnh mạn tính như cao huyết áp, tiểu đường… được chỉ định điều trị bằng 10 loại thuốc hóa dược hoặc hơn thế nữa. Họ bị các phản ứng phụ gây ho, viêm đường hô hấp trên, đau nhức cơ bắp, giảm trí nhớ và cả suy thận.
Khi người bệnh hỏi bác sỹ của họ có cách nào thoát khỏi những đau đớn, khổ ải về bệnh tật và thuốc men… câu trả lời thường là không. Với họ chỉ có con đường duy nhất là dùng thêm thuốc để chữa trị các triệu chứng do thuốc gây ra và đi chạy thận nhân tạo.
Các bác sỹ ít khi nói tới khả năng hỗ trợ điều trị rất hiệu quả bằng cách thay đổi chế độ ăn uống, cách luyện tập và sinh hoạt, các phương pháp dưỡng sinh, thiền, thư giãn… cũng như dùng nhiều chất có nguồn gốc thiên nhiên như: nhiều loại thảo dược, vitamin như D, K, dầu cá và các loại thức ăn có nhiều acid béo omega 3, chất khoáng ma-nhê và nếu xa hơn nữa là các thực phẩm chức năng như: Alpha lipoic acid, L-carnitine, Taurine, các sản phẩm trợ sinh (probiotic)…
Những chất này đã được nghiên cứu và cho thấy là có tác dụng phòng và chữa nhiều bệnh mạn tính và không có tác dụng phụ nguy hiểm. Các sản phẩm này chỉ có một “tội” là: không thể bảo vệ được bản quyền và không thể mang lại lợi nhuận kéo dài trong hơn 20 năm cho hãng dược phẩm muốn bào chế, thử nghiệm và đưa chúng vào thị trường như một sản phẩm có tác dụng phòng và chữa bệnh.
Một điều chúng ta cần nhớ là ở giai đoạn cuối những năm 30 thế kỷ trước, có tới 80% các dược phẩm được bào chế và ứng dụng có nguồn gốc thiên nhiên xuất phát từ các kinh nghiệm y học dân gian và ngay hiện tại có tới 25% các dược phẩm đang lưu hành được chiết xuất từ thảo dược.
Theo thống kê của Tổ chức Y tế Thế giới, ngày nay các thảo dược vẫn được sử dụng như những phương tiện phòng và chữa bệnh chính cho hơn 80% nhân loại. Vì các lý do kinh tế, các hãng dược không muốn phát triển và sử dụng thuốc thiên nhiên dưới dạng nguyên sinh.
Việc tách ra một hợp chất có tác dụng vào một cơ chế trong bệnh lý cùng việc biến đổi các phân tử tự nhiên bằng phương pháp công nghệ hóa, công nghệ gien để có thể sở hữu bản quyền bởi các hãng dược phẩm trong rất nhiều trường hợp đã làm giảm hiệu quả cho một dược phẩm có nguồn gốc thiên nhiên.
Tại sao cần kết hợp các sản phẩm có nguồn gốc thiên nhiên trong phòng và điều trị các bệnh cấp và mạn tính?
Nhiều bệnh cấp tính như cảm, cúm, viêm nhiễm do vi trùng, siêu vi trùng, nấm bệnh… xuất hiện khi chúng ta không được khỏe, do hệ miễn dịch của chúng ta suy yếu. Các bệnh mạn tính phổ biến và gây tử vong nhiều nhất cho con người như bệnh tim mạch, ung thư, tiểu đường, tắc nghẽn phổi, hen suyễn, béo phì, trầm cảm, các hội chứng chuyển hóa và rối loạn hệ tiêu hóa, bệnh viêm khớp, dị ứng, u nang, u xơ… phần nhiều có nguồn gốc từ chế độ dinh dưỡng lệch lạc, môi trường ô nhiễm và cuộc sống tinh thần căng thẳng…
Các thuốc hóa dược đại bộ phận dùng để làm giảm triệu chứng, làm tăng chỉ số này, giảm chỉ số kia trong cơ thể người bệnh, nhưng rất ít khi chúng động tới nguyên nhân của bệnh tật và vì thế hầu như không có khả năng chữa khỏi bệnh. Các thuốc hóa dược đều có những hậu quả phụ và nhiều khi các phản ứng này được nhận thức và điều trị bằng một thuốc khác như một bệnh lý mới.
Thực tế đã cho thấy nhiều thuốc men có thể được thay thế bằng vitamin, dược thảo, axit amin, sản phẩm trợ sinh, thức ăn sạch và giàu chất dinh dưỡng, cả thực phẩm chức năng cũng như biện pháp không dùng thuốc.
Việc người ta khẳng định chỉ có thuốc do các hãng dược bào chế và được bác sỹ kê đơn mới có tác dụng phòng và chữa bệnh chẳng qua là việc làm theo định kiến và sự lệ thuộc kinh tế vào công nghệ y dược hiện đại.
Vì sao thực phẩm chức năng và các phương pháp điều trị không dùng thuốc men ít được nghiên cứu, được phổ biến để ứng dụng rộng rãi?
Con người, với trí tuệ, kinh nghiệm và khả năng quan sát, phân tích của họ, đã tìm ra và ứng dụng rất nhiều sản phẩm có nguồn gốc thiên nhiên để nâng cao sức khỏe, chất lượng cuộc sống, phòng và chữa bệnh. Bản thân con người, cùng những thứ cần thiết nhất để chúng ta sống và tồn tại, phát triển, làm nên những việc phi thường, đều do thiên nhiên tạo ra.
Khoa học chỉ góp phần thúc đẩy sự phát triển và nâng cao đời sống của chúng ta dựa vào những quy luật tự nhiên và những gì có trong thiên nhiên. Bởi vậy, đừng nên lấy khoa học làm con “ngáo ộp” và những tôn chỉ của tổ chức này, đại học kia… để làm thước đo và luật lệ cho mọi vấn đề, nhất là khi vấn đề đó liên quan tới sức khỏe và bệnh tật.
Mặc dù có những tiến bộ nhất định, nhưng những hiểu biết về cơ thể con người, bệnh tật cũng như cách phòng và điều trị của y học ngày nay còn rất hạn chế. Hiểu biết của chúng ta về khả năng phòng và chữa bệnh của các chất có nguồn gốc thiên nhiên cũng như sức mạnh tiềm tàng của cơ thể hiện nay chỉ là một con số giới hạn của sự vô hạn.
Nhưng cách nói như “không thể làm được” cái gì “vì khoa học chưa chứng minh được”, là sự vô trách nhiệm phổ biến của những người lầm tưởng mình đang đứng ở đỉnh cao của khoa học.
Một điều đáng tiếc là đã có rất nhiều nghiên cứu cơ bản, nghiên cứu dịch tễ học và lâm sàng cho thấy khả năng phòng và chữa bệnh ưu việt của nhiều sản phẩm thiên nhiên, thực phẩm chức năng và cả phương pháp chữa bệnh không dùng thuốc, đã bị vùi dập vào dĩ vãng.
Các tác giả nghiên cứu rất ít khi công bố được công trình của họ trên các tạp chí y khoa lớn vì đại bộ phận thẩm định viên của các tạp chí này thiên về khuynh hướng y dược hiện đại.
Các nguồn tài trợ, kinh phí để tiếp tục nghiên cứu và ứng dụng các sản phẩm hoặc phương pháp này cũng rất hạn chế để có thể cạnh tranh được với số tiền lên tới hàng tỉ đô-la mà các hãng dược phẩm đa quốc gia có thể bỏ ra để làm thuốc và quảng bá chúng. Việt Nam có đủ kinh nghiệm và tiềm năng để trở thành cường quốc về y học thiên nhiên.
Với đường lối đúng đắn, sự học hỏi thường xuyên và vận dụng sáng tạo, y học dân tộc và thiên nhiên Việt Nam đã đặt được nền móng ở hầu hết các bệnh viện, trạm xá, các đại học y… với những bài thuốc và phương pháp chữa bệnh rất độc đáo và hiệu quả.
Một ví dụ có thể đơn cử ở đây là sự phát hiện và ứng dụng Artemisinin và các dẫn xuất để điều trị bệnh sốt rét. Dược phẩm này đã được chiết xuất từ cây thanh hao hoa vàng bởi các nhà khoa học Trung Quốc, Mỹ và đã được ứng dụng để điều trị cũng như cứu mạng hàng triệu người bệnh ở châu Phi và châu Á.
Artemisinin đã được phát hiện nhờ những tài liệu cổ điển có từ hàng nghìn năm trước và hầu như không làm đau một súc vật thực nghiệm nào cũng như không cần chi phí tốn kém cho các thử nghiệm lâm sàng. Mặc dù kết quả điều trị cũng như độ an toàn của Artemisinin theo kinh nghiệm của các bác sỹ ở Việt Nam và châu Phi là vượt trội so với các thuốc cổ điển, Cơ quan quản lý về dược phẩm của Hoa Kỳ (FDA) đã áp đặt và kìm hãm việc sử dụng Artemisinin trong một thời gian dài chỉ với lý do: Nó chưa được kiểm nghiệm qua thử nghiệm lâm sàng.
Đến khi các nước châu Phi đồng loạt phản đối và không tuân theo sự phán quyết của FDA và Chính phủ Mỹ, thì Tổ chức Y tế Thế giới đã phải chấp nhận đưa Artemisinin vào các chương trình phòng chống và điều trị sốt rét trên toàn cầu.
Những nhà khoa học tham gia vào phát hiện và tách lọc được Artemisinin đáng nhẽ phải xứng đáng được 2 giải Nobel về y học, bởi vì họ đã làm một việc mang lại lợi ích cho những bệnh nhân bị sốt rét (đại bộ phận là người nghèo) to lớn hơn nhiều so với các công trình khoa học và phát minh đã được giải Nobel trong những năm gần đây.
Các sản phẩm thiên nhiên và thực phẩm chức năng có đáng sợ không?
Nếu được bào chế đúng tiêu chuẩn và dùng đúng, hầu hết các sản phẩm này có tính an toàn vượt trội so với các dược phẩm. Chúng ta phải thấy vui mừng vì nước nhà đã có những hãng bào chế và phân phối có uy tín, trách nhiệm với sản phẩm thuốc thiên nhiên, cũng như thực phẩm chức năng có chất lượng và giá trị phòng, chữa bệnh tốt. Mặc dù những nguy cơ, tác hại của thuốc thiên nhiên cũng như thực phẩm chức năng luôn được đề cập và thổi phồng, nhưng những con số sau đây nói lên tất cả:
– Năm 2005: Không có một trường hợp tử vong nào do vitamin và thực phẩm chức năng được báo cáo ở Mỹ, mặc dù có tới hơn 70% người Mỹ dùng các sản phẩm này (theo Trung tâm quản lý về ngộ độc của Mỹ, trang 588 KBPDF).
– Con số tử vong do dùng thuốc theo đúng chỉ định gây ra trong các bệnh viện tại Mỹ năm 2005 là khoảng 106 nghìn trường hợp (theo Tạp chí của Hiệp hội Y khoa Mỹ).
Trong lĩnh vực nào của đời sống cũng có kẻ xấu người tốt. Chúng ta hãy giúp người dân hiểu biết và loại bỏ những sản phẩm tồi, nhưng đồng thời cần tạo điều kiện giúp đỡ, phát triển những phát hiện và ứng dụng của thực phẩm chức năng, thuốc thiên nhiên mới và cổ điển vì lợi ích của người bệnh và cả nền kinh tế của đất nước.
Nếu làm được việc này, Y tế Việt Nam có thể đón trước chiều hướng không thể thay đổi của nhân loại trong một tương lai không xa, đó là quay lại với các sản phẩm thiên nhiên để phòng và chữa bệnh.
Nếu biết lắng nghe, học hỏi, tiếp thu kiến thức dân gian và luôn tìm kiếm, chúng ta có thể sẽ tìm được cách khống chế và hạn chế những căn bệnh hiểm nghèo nhất của thời đại. Như Paracelsus, một triết gia vĩ đại của nhân loại, đã nói: “Thiên nhiên không tạo ra bệnh gì mà không có thuốc để trị nó”.
TS.BS. Hoàng Xuân Ba
Tiến sỹ Hoàng Xuân Ba là một huyền thoại về y học chữa Ung thư tại Mỹ. Ông là người đưa ra phương pháp ” bỏ đói tế bào ung thư ” và “ngăn chặn cơ chế tạo mạch” , khác với phương pháp “xạ trị tế bào ung thư” thông thường.
Một video rất ấn tượng của tiến sỹ William Li thuyết trình về việc bỏ đói tế bào ung thư và ngăn chặn cơ chế tạo mạch
Tiếp theo thông tin từ huynh Hạt Gạo Làng, tại hạ cũng có một số điều rút ra từ buổi nói chuyện đó muốn chia sẻ với mọi người như sau:
Nguyên lý điều trị của BS.TS Hoàng Xuân Ba là dùng thảo dược và các chất dinh dưỡng để giúp cơ thể tự điều chỉnh lại sự kích thích màng tế bào và chuyển hóa năng lượng. Các chế phẩm này không trực tiếp tiêu diệt khối u mà giúp màng tế bào ổn định, không bị kích thích cũng như điều hòa hoạt động chuyển hóa và tổng hợp năng lượng của tế bào, đào thải gốc oxi hóa tự do, ngăn chặn quá trình dị sản của các tế bào nhờ đó bệnh nhân có thể sống lâu hơn với chất lượng cuộc sống tốt hơn…. Thực tế, nhiều bệnh nhân sau khi được điều trị, kết quả chiếu chụp và xét nghiệm cho thấy khối u bị tiêu mất hoàn toàn hoặc bị vôi hóa, sẹo hóa.
Nhiều người lầm tưởng phương pháp BS TS Hoàng Xuân Ba đơn thuần là Đông y, nhưng không phải. Gốc rễ vấn đề vẫn là khoa học, là có sự nghiên cứu và hiểu biết sâu rộng về Tây y, chỉ khác người khác là BS Ba không những giỏi về Hóa, Sinh mà còn rất giỏi về Toán và Lý. Chính vì thế, BS TS Hoàng Xuân Ba hiểu rất rõ cơ chế các bệnh và tính lý học của từng người bệnh. Từ sự hiểu biết sâu sắc khoa học hiện đại, BS Ba tìm những hoạt chất chữa được bệnh trong thiên nhiên từ thảo dược, cỏ cây, .. mà không dùng hóa dược. Chính vì thế mà phương pháp của BS Ba trong một loạt bệnh mãn tính, nan y không những đạt hiệu quả rất cao mà còn an toàn, không hề có tác dụng phụ.
Vài dòng ghi nhận từ Buổi nói chuyện 17-3
Đối với bệnh gai cột sống và thoát vị đĩa đệm, tôi có những thông tin sau hy vọng giúp được bạn:
Đối với bệnh này tuy chữa được, không còn đau đớn nữa nhưng cũng không khỏi hẳn, vẫn phải kiêng khem và tránh mang vác nặng.
Theo phương pháp chữa này, đối với những bệnh nhẹ thì chỉ cần uống thuốc, nhưng với những bệnh mãn tính thì cần phải truyền thuốc trực tiếp vào cơ thể. Trước mắt bạn có thể uống thuốc Flamasol và Artemune. Sau đó, bạn phải ra Hà Nội truyền thuốc do bác sĩ Ba trực tiếp pha cho bệnh nhân và truyền vào cột sống. Do thuốc này không để được lâu nên chỉ khi nào bắt đầu truyền mới pha thuốc để truyền.
Thời lượng truyền phải đạt được 60 lần truyền trong thời gian 3 tháng thì mới ổn định, cứ 1 tháng truyền 20 ngày và cứ truyền được 5 ngày thì nghỉ 2 ngày.Sau đợt truyền thuốc này thì chỉ cần uống thuốc chứ không phải đi truyền lại nữa.
Sau đây là những công dụng của thuốc uống cho bạn và những người quan tâm tham khảo:
Flamasol
Hỗ trợ điều trị các bệnh lý viêm khớp, đau khớp, thoái hoá khớp và một số bệnh lý khác
– Công dụng: Hỗ trợ điều trị viêm đau khớp: thấp khớp, viêm khớp dạng thấp, viêm khớp vảy nến, thoái hoá khớp.Hỗ trợ điều trị cho các bệnh viêm mãn tính như viêm đại tràng mãn tính, viêm mạch máu tự miễn, lupus đỏ…
– Thành phần: Nhũ hương; Nhàu; Bạch thược; Thổ phục linh; Hoàng bá; Cây sói rừng; L-carnitine fumarate.
Nhũ hương: có tác dụng chống viêm, sử dụng nhiều cho bệnh viêm mãn tính, đặc biệt là viêm khớp, thoái hoá khớp.
Nhàu: quả Nhàu chứa nhiều acid linoleci có tác dụng chống viêm nhiễm rất tốt. Chiết xuất cây Nhàu còn được sử dụng làm chất bổ dưỡng, có tác dụng chữa các bệnh về mắt, da, nướu răng, họng, đau dạ dày và táo bón.
Bạch thược: có tác dụng chống viêm và hạ nhiệt, chống sự hình thành huyết khối do tiểu cầu, tăng lưu lượng máu nuôi cơ tim, có tác dụng hạ men gan, bảo vệ gan. Bạch thược còn dùng chữa đau bụng do co thắt, lưng ngực đau, chân tay nhức mỏi…
Thổ phục linh: có tác dụng khử độc, mạnh gân xương, tán uất kết. Thổ phục linh được sử dụng để chữa trị các bệnh như: rối loạn tiêu hoá, viêm bàng quang, viêm khớp, chân thương ngoài da…
Hoàng bá: có tác dụng kháng khuẩn, kháng nấm, chống viêm, ức chế miễn dịch.
Cây Sói rừng – Sarcandra glabra: có tác dụng thanh nhiệt, giải độc, tăng sự lưu thông tuần hoàn, giảm đau, kháng khuẩn và điều hoà miễn dịch. Cây Sói rừng được sử dụng trong các bệnh lý viêm, nhiễm khuẩn, các bệnh tự miễn, viêm khớp dạng thấp…
L-carnitine Fumarate: đóng vai trò quan trọng trong việc sử dụng và biến chất béo thành năng.
Artemune
Có tác dụng thanh nhiệt, tăng cường chức năng thải độc cơ thể. Sử dụng tốt trong trường hợp da nổi mụn do chức năng Gan kém, sau hoặc trong thời gian sử dụng kháng sinh. Artemune có chức năng tăng cường hệ miễn dịch cơ thể.
Các trường hợp sử dụng Artemune tốt nhất:
– Da bị nổi mụn do nóng trong.
– Sử dụng kháng sinh dài ngày
– Chức năng thải độc của Gan suy giảm
– Phòng ngừa bị viêm Gan
– Những người sử dụng nhiều rượu bia, đồ uống có cồn.
– Trường hợp bị dị ứng, mẩn ngứa do chức năng Gan kém sử dụng liền trong 3 tháng để có kết quả tốt nhất
Chi tiết sản phảm:
Thành phần: Thanh hao hoa vàng, Nhân trần bắc, Sói rừng, Hoàng kỳ, Hoài sơn, Succinic Acid, Pregnenolone, Phụ liệu vừa đủ 1 viên.
Công dụng: Giúp thanh nhiệt, lợi mật, tăng cường chức năng giải độc, hỗ trợ các bệnh viêm gan, viêm khớp dạng thấp, bệnh tự miễn, điều hoà chức năng miễn dịch. Điều hòa lượng và cân bằng kiềm toan trong cơ thể.
Thân mến
Bài viết trước Thanh Tước đã post lên một số tài liệu về bệnh hen suyễn, xoang , viêm mũi dị ứng để các bạn tìm hiểu triệu chứng của bệnh hen suyễn và tìm hiểu cách điều trị của Bác sỹ Tiến sỹ Hoàng Xuân Ba. Anh Lệnh Hồ Xung đã bổ sung thêm thông tin cho hình ảnh mà Thanh Tước nêu, Allergy Research Group chính là nơi làm việc của Dr Ba Hoàng bên Mỹ. Bs.TS Hoàng Xuân Ba là Giám đốc nghiên cứu khoa học, phụ trách toàn bộ về nghiên cứu Miễn dịch của Viện nghiên cứu miễn dịch Hoa Kỳ (Allergy Research Group. Nhóm các nhà khoa học này đã nghiên cứu để hiểu về bệnh lý và tìm cách điều trị bệnh suyễn một cách hiệu quả và an toàn. Kết quả hơn 10 năm nghiên cứu đã mang lại sự thành công của sản phẩm Phytocort với những thảo dược lành tính bổ dưỡng nhưng có kết quả chữa bệnh hơn rất nhiều so với thuốc tây( kháng sinh). Vì thế Thanh tước xin chân thành khuyên các bạn nên sử dụng sản phẩm được chiết xuất bào chế từ thảo dược mà Tiểu Phương và Anh Thế Anh vẫn thường chia sẽ cùng các bạn khi trị các căn bệnh nan y hay các bệnh mãn tính.
Thanh Tước xin trích lại bài viết của anh Thế Anh về Bệnh hen suyễn, xoang, viêm mũi dị ứng. Phác đồ cho bệnh tai mũi họng và liều lượng dùng có thể áp dụng như sau – cho bệnh nhân chưa tới giai đoạn mạn tính, hoặc quá muộn:
1.Pharysol
2.Panax Co
3.Thuốc xịt
Thanh Tước xin giải thích thêm Sản phẩm Phytocort trong tài liệu Thanh Tước gửi các bạn là công trình nghiên cứu của Bác sỹ Ba nhưng tại sao không có trong phác đồ . Bởi vì Bác sỹ Ba đã nghiên cứu ra 1 sẩn phẩm mới hữu hiệu và an toàn hơn nữa đó chính là PANAX CO thay thế Phytocort và Pumasol.
Nếu các bạn bị bệnh hô hấp bình thường chỉ cần dùng pharysol va panax co là đủ rồi, nếu ho viêm họng cấp mãn tính có đờm thì bên cạnh Pharysol và Panax Co thì nên dùng thêm futasol
Ngoài ra khi các bạn bị viêm xoang mà đau răng nhức óc thì bên cạnh pharysol và Panax co dùng thêm Tomonan, tuy nhiên có 1 vài trường hợp phải thêm thuốc khác Ví dụ hen suyễn thì các bạn dùng Pharysol , panax co và thêm Phytosaponin.
Còn bị Amidan bình thường không nặng lắm các bạn chỉ cần dùng Pharysol, Hemican, nếu Amidan mà nặng bên cạnh Pharysol, Hemican cần phải dùng thêm Panax Co.
Trên đây là những chia sẽ của Thanh tước tới tất cả các bạn!
Chúc các bạn và gia đình luôn mạnh khỏe!
Việc sử dụng thuốc tây ( kháng sinh) thực sự rất vô cùng nguy hại cho cơ thể, rất dễ làm hư thận là đúng. Thanh Tước có người bạn vào bệnh viện vì bệnh hen suyễn uống thuốc tây mà sau đợt đều trị đã tăng hơn 3kg khuôn mặt phù nề trông khiếp lắm do bị ứ nước trong thận rất khó bài tiết ra ngoài. Như vậy chức năng thận đã hoạt động kém đi sau lần điều trị và chắc chắn chỉ mấy lần vào viện như vậy nữa coi như hỏng luôn thận. Để các bạn và người thân trong gia đình tránh xa thuốc tây thì Thanh Tước nghĩ các bạn hãy thường xuyên vào trang Y hoc và sức khỏe các bạn có thể gặp anh Thế Anh tư vấn cho các bạn nhiều loại bệnh nhiều phác đồ điều trị và chỉ cần sử dụng thuốc theo phác đồ mà do bác sỹ Ba kê đảm bảo sức khỏe của các bạn và người thân sẽ rất tốt.
Anh Lệnh Hồ Xung bảo sẽ tìm lại một số bài về bệnh hen suyễn post lên cho các ban thao khảo nhưng chắc anh bận chưa tìm được . Hiện tại Thanh Tước tìm được tài liệu nên xin posrt dùm anh Lệnh Hồ Xung để các bạn tìm hiểu cách chữa bệnh theo phương pháp của bác sỹ Hoàng Xuân Ba mà theo Thanh Tước biết thì trước khi là bác sỹ chữa bệnh ung thư và các bệnh nan y nổi tiếng thì bác sỹ Ba nổi tiếng chữa bệnh hen suyễn và chữa xoang.
Mới đây sản phẩm Thảo mộc Hương lại vừa cho ra đời một sản phẩm mới Panaxco để chữa bệnh xoang và hen suyễn thay thế Pulmasol mà Bác sỹ Ba đã dày công nghiên cứu và được đánh giá rất hữu hiệu an toàn hơn hẳn thuốc cũ. mình post Tài lệu để các bạn có thể tự điều trị bệnh cho chính mình và những người thân của mình nhé.




